The equations of Hodgkin and Huxley provide a good description of the electro-physiological properties of the giant axon of the squid. These equations capture the essence of spike generation by sodium and potassium ion channels. The basic mechanism of generating action potentials is a short influx of sodium ions that is followed by an efflux of potassium ions. This mechanism is essentially preserved in higher organisms. Cortical neurons in vertebrates, however, exhibit a much richer repertoire of electro-physiological properties than the squid axon studied by Hodgkin and Huxley. These properties are mostly due to a large variety of different ion channels (Hille, 1992; Llinás, 1988; Koch, 1999).
In this section we give an overview of some of the ion channels encountered in different neurons. The basic equation of detailed neuron models is more or less the same as that of the Hodgkin-Huxley model (Hodgkin and Huxley, 1952) except that it contains more types of ion channels. The membrane potential u of the neuron is given by
| C |
(2.9) |
| Ik = |
(2.10) |
![\begin{center}
\begin{minipage}{0.8\textwidth}
{\bf A}\\
\includegraphics[wi...
...\includegraphics[width=0.5\textwidth]{tau_IT.ps.gz}
\end{minipage} \end{center}](img129.gif) |
Apart from fast sodium ion channels, which are qualitatively similar to those of the Hodgkin-Huxley model and denoted by INa, some neurons contain a `persistent' or `non-inactivating' sodium current INaP. `Non-inactivating' means that this current has qNaP = 0. In other words, it is described by an activation variable m only and does not have a separate inactivation variable h,
| INaP = |
(2.11) |
Voltage clamp experiments on various neurons have revealed that there is more
than just one type of potassium channel. In thalamic relay neurons, for
example, two different types of potassium channels are present, viz. a
rapidly inactivating potassium current
IA (inactivation time
constant
![]()
![]() 10 ms) and a slowly inactivating potassium current
IK2 (time constant
10 ms) and a slowly inactivating potassium current
IK2 (time constant
![]()
![]() 200...2000 ms); cf. Fig. 2.9. Both classes can even be further subdivided into two
different subtypes
IA1,
IA2, and
IK2a,
IK2b, respectively, in order to fit
activation and inactivation curves that contain more than a single time
constant (Huguenard and McCormick, 1992).
200...2000 ms); cf. Fig. 2.9. Both classes can even be further subdivided into two
different subtypes
IA1,
IA2, and
IK2a,
IK2b, respectively, in order to fit
activation and inactivation curves that contain more than a single time
constant (Huguenard and McCormick, 1992).
![\begin{center}
\begin{minipage}{0.8\textwidth}
\par {\bf A}\\
\includegraphic...
...ludegraphics[width=0.5\textwidth]{tau_IK2.ps.gz}
\par\end{minipage} \end{center}](img133.gif) |
The afore mentioned A-current IA is just one member of a large class of transient potassium currents. These currents have in common an inactivation time constant of a few tens of milliseconds which is rather long as compared to the sodium dynamics. Since potassium currents are outward currents, the current IA tends to hyperpolarize the membrane potential and therefore slows down the firing of action potentials. For weak sustained stimulation, new action potentials only occur after the A-current has become sufficiently inactivated (Connor et al., 1977).
In a similar way, A-currents are responsible for a long delay between the sudden onset of an excitatory stimulus and the very first action potential; cf. Fig. 2.10. The onset of a depolarizing current will quickly lead to an activation of the A-current which in turn counteracts a depolarization of the membrane potential. Following the slow inactivation of the A-current, the membrane potential finally rises to the firing threshold and an action potential is released.
![\begin{minipage}[t]{0.45\textwidth}
{\bf A}\\
\includegraphics[clip,width=\te...
...B}\\
\includegraphics[clip,width=\textwidth]{low_freq_b.ps.gz}
\end{minipage}](img134.gif) |
The gain function of the Hodgkin-Huxley model is discontinuous at the firing threshold which means that it cannot produce low-frequency spike trains with a constant input current. Nevertheless, low firing rates can also be achieved without the inactivating current IA provided that the potassium currents have suitable characteristics. In a model of neocortical interneurons (Erisir et al., 1999), there are two non-inactivating potassium channels and an inactivating sodium channel of the Hodgkin-Huxley type. With a suitable choice of parameters, the gain function is continuous, i.e., repetitive firing is possible in the full range of frequencies between zero and a few hundred Hertz; cf. Fig. 2.11B. A short current pulse from rest can elicit action potentials if a critical threshold voltage is exceeded. In contrast to the Hodgkin-Huxley model, stimulation that is only slightly above threshold leads to delayed pulse generation; cf. Fig. 2.11B. Neurons with continuous gain function and delayed pulse generation are called type I whereas neurons with discontinuous gain function are called type II. The distinction between type I and type II plays an important role in Chapter 3.
![\hbox{{\bf A} \hspace{70mm} {\bf B}}
\hbox{\hspace{10mm}
\includegraphics[widt...
...0mm}
\includegraphics[width=50mm]{Figs-ch-detailed-models/A0pulses-all.eps}
}](img135.gif) |
Apart from channels for sodium and potassium ions, many central neurons
contain also channels for calcium ions. Similarly to the common sodium and
potassium currents, calcium currents are described in terms of a maximum
permeability ![]() times a combination of activation m and
inactivation h variables. In contrast to sodium and potassium the
intracellular calcium concentration is very low and can be significantly
affected by the calcium influx during an action potential. The current that
passes through open channels thus depends on the actual calcium concentrations
[Ca2+]i, e inside and outside the cell, and on the
voltage u across the membrane. The conductivity of a calcium channel
is described by the Goldman-Hodgkin-Katz equation which accounts for a small
nonlinearity in the voltage/current-relation of the open channel that
is caused by the large concentration gradient across the membrane
(Hille, 1992, chapt. 13). Altogether, the current through calcium channel
x is given by
times a combination of activation m and
inactivation h variables. In contrast to sodium and potassium the
intracellular calcium concentration is very low and can be significantly
affected by the calcium influx during an action potential. The current that
passes through open channels thus depends on the actual calcium concentrations
[Ca2+]i, e inside and outside the cell, and on the
voltage u across the membrane. The conductivity of a calcium channel
is described by the Goldman-Hodgkin-Katz equation which accounts for a small
nonlinearity in the voltage/current-relation of the open channel that
is caused by the large concentration gradient across the membrane
(Hille, 1992, chapt. 13). Altogether, the current through calcium channel
x is given by
Neurons of the deep cerebellar nuclei, for example, contain two different types of calcium channels that give rise to a so-called high-threshold calcium current ( IL) and a low-threshold calcium current ( IT), respectively (Llinás and Mühlethaler, 1988; Jahnsen, 1986). From a functional point of view, the low-threshold calcium current is particularly interesting, because it is responsible for a phenomenon called post-inhibitory rebound. Post-inhibitory rebound means that a hyperpolarizing current, which is suddenly switched off, results in an overshoot of the membrane potential or even in the triggering of one or more action potentials. Through this mechanism, action potentials can be triggered by inhibitory input. These action potentials, however, occur with a certain delay after the arrival of the inhibitory input, viz., after the end of the IPSP (Aizenman and Linden, 1999).
The low-threshold calcium current is `inactivating', i.e., the corresponding
ion channel is shutting down after the membrane has become depolarized (qx = 1). In this respect, the
IT current is similar to the fast
sodium current of the Hodgkin-Huxley model. The discerning feature of the
IT current, however, is the fact that the activation and
inactivation curves are shifted significantly towards a hyperpolarized
membrane potential so that the channel is completely inactivated (
h ![]() 0) at the resting potential; see Fig. 2.8C. This is
qualitatively different from the situation of the sodium channels of the
Hodgkin-Huxley model. These channels are also closed at rest; the reason,
however, is different. In the Hodgkin-Huxley model the sodium channels are
not activated (
m
0) at the resting potential; see Fig. 2.8C. This is
qualitatively different from the situation of the sodium channels of the
Hodgkin-Huxley model. These channels are also closed at rest; the reason,
however, is different. In the Hodgkin-Huxley model the sodium channels are
not activated (
m ![]() 0) whereas the T-type calcium current is
inactivated (
h
0) whereas the T-type calcium current is
inactivated (
h ![]() 0); cf. Fig. 2.8B and C.
0); cf. Fig. 2.8B and C.
The basic mechanism of post-inhibitory rebound can be easily understood by means of Fig. 2.8C which shows the stationary values of the activation and the inactivation variables, together with their time constants as a function of the membrane potential. In order to open the T-type calcium channels it is first of all necessary to remove its inactivation by hyperpolarizing the membrane. The time constant of the inactivation variable h is rather high (dashed line in the right panel of Fig. 2.8C) and it thus takes a while until h has reached a value sufficiently above zero. But even if the channels have been successfully `de-inactivated' they remain in a closed state, because the activation variable m is zero as long as the membrane is hyperpolarized; cf. left panel of Fig. 2.8C. However, the channels will be transiently opened if the membrane potential is rapidly relaxed from the hyperpolarized level to the resting potential, because activation is faster than inactivation and, thus, there is a short period when both m and h are non-zero. The current that passes through the channels is terminated (`inactivated') as soon as the inactivation variable h has dropped to zero again. The resulting current pulse is called a low-threshold calcium spike.
The increase in the membrane potential caused by the low-threshold calcium spike may be sufficient to trigger ordinary sodium action potentials. We will refer to action potentials generated by this mechanism as rebound spikes. Figure 2.12A shows an example of rebound spikes that are triggered by an inhibitory current pulse. The time course of the activation and inactivation variables of the low-threshold calcium-current are plotted in Fig. 2.12B.
![\begin{minipage}[t]{0.45\textwidth}
{\bf A}\\
\includegraphics[clip,width=\te...
... {\bf B}\\
\includegraphics[clip,width=\textwidth]{dcn2.ps.gz}
\end{minipage}](img140.gif) |
![\begin{center}
\begin{minipage}{0.8\textwidth}
\par % \{ bf A\}
\includegraph...
...ncludegraphics[width=0.5 textwidth]\{tau_Ih.ps.gz\}
\end{minipage} \end{center}](img141.gif) |
In addition to the low-threshold calcium current, some neurons, e.g., cortical pyramidal cells, contain a second type of calcium current, called high-threshold calcium current, or IL-current for short. Similar to the persistent sodium current this current is non-inactivating (or `long-lasting' - hence the index L), but it is activated only at rather high levels of depolarization; cf. Fig. 2.13A. The IL-channels are thus open during action potentials, but otherwise closed. The current through these channels is described by the Goldman-Hodgkin-Katz equation Eq. (2.12) with px = 2 and qx = 0, i.e., the inactivation variable h is absent.
The role of calcium for generating spikes is twofold. On the one hand, calcium ions carry a positive electrical charge and contribute therefore to the depolarization of the membrane. On the other hand, calcium ions are an important second messenger that is involved in all kinds of intracellular signaling. An example of the latter role of calcium ions is provided by Ca2+-activated potassium channels. The potassium current IC that passes through these channels is described by the familiar equation
| IC = |
(2.13) |
| (2.14) |
| (2.15) |
![\begin{center}
\begin{minipage}{0.4\textwidth}
{\bf A}\\
\includegraphics[wi...
...
\includegraphics[width=\textwidth]{tau_IC.ps.gz}
\end{minipage} \end{center}](img148.gif) |
The plot in Fig. 2.14 shows that the stationary value of
m is growing with increasing calcium concentration. Similarly, the
relaxation towards the stationary value becomes faster since the time
constant ![]() decreases with increasing calcium concentration;
cf. Fig. 2.14B. The
IC current is thus activated by intracellular calcium, as indicated by its name. Due
to the short time constant of its activation variable the maximum of
this potassium current is reached during or immediately after an
action potential. If present, the calcium-activated potassium channels
can provide a major contribution to the repolarization of the membrane
after an action potential.
decreases with increasing calcium concentration;
cf. Fig. 2.14B. The
IC current is thus activated by intracellular calcium, as indicated by its name. Due
to the short time constant of its activation variable the maximum of
this potassium current is reached during or immediately after an
action potential. If present, the calcium-activated potassium channels
can provide a major contribution to the repolarization of the membrane
after an action potential.
Closely related to the IC-current is another Ca2+-activated potassium current, the so-called IAHP-current (AHP is short for afterhyperpolarization). In contrast to its C-type sibling the activation of this current is much slower and and purely [Ca2+]i-dependent, i.e., voltage independent. This current is activated by calcium ions that enter through (voltage-gated) L-type channels during an action potential (Tanabe et al., 1998). More precisely, we have
| IAHP = |
(2.16) |
| (2.17) |
Because of the fact that the AHP-channels are not inactivating and because of their low time constant at rest, each action potential simply increases the activation m by a more or less fixed amount. If the neuron is stimulated by a constant depolarizing current each action potential increases the amount of open AHP-channels and the corresponding potassium current subtracts from the applied stimulus. The firing frequency is thus decreasing, a phenomenon that is known as firing frequency adaptation; cf. Fig. 2.15.
![\begin{minipage}[t]{0.45\textwidth}
{\bf A}\\
\includegraphics[clip,height=0....
...hics[clip,height=0.6\textwidth,trim=0 5 0 6]{adaptation_b.ps.gz}
\end{minipage}](img152.gif) |
Among all ions of the intra- and extra-cellular fluid calcium ions are particularly interesting. On the one hand, they contribute to the ionic current across the membrane; see Section 2.3.3. On the other hand, some potassium channels are controlled by the intracellular calcium concentration; see Section 2.3.4. These channels are not present in the model of Hodgkin and Huxley, but they are responsible for interesting electro-physiological properties of neurons in different brain areas.
Whereas sodium and potassium concentrations are large as compared to the ionic in- or efflux during an action potential, the intracellular calcium concentration is not and, hence, it cannot be treated as a constant. Since both the influx through calcium channels and the state of the afore mentioned potassium channels depend on the intracellular calcium concentration we have to describe this concentration explicitly by a variable [Ca2+]i.
Modeling the calcium concentration, however, is no trivial task because both diffusion and chemical calcium buffers ought to be taken into account. To avoid these difficulties a purely phenomenological approach can be adopted. The intracellular calcium concentration [Ca2+]i is described by a linear differential equation,
![$\displaystyle {\frac{{{\text{d}}[\text{Ca}^{2+}]_{\text{i}}}}{{{\text{d}}t}}}$](img153.gif) = - = - |
(2.18) |
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
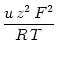
![$\displaystyle {\frac{{[\text{Ca}^{2+}]_{\text{e}} \, {\text{e}}^{-u\,z\,F/R\,T} - [\text{Ca}^{2+}]_{\text{i}}}}{{1-{\text{e}}^{-u\,z\,F/R\,T}}}}$](img139.gif) ,
,