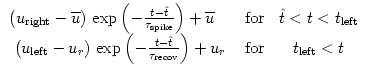We have seen in the previous chapter the Hodgkin-Huxley model does not have a clear-cut firing threshold. Nevertheless, there is a critical regime where the sensitivity to input current pulses is so high that it can be fairly well approximated by a threshold. For weak stimuli, the voltage trace returns more or less directly to the resting potentials. For stronger stimuli it makes a large detour, that is, emits a spike; see Fig. 3.10B. This property is characteristic for a large class of systems collectively termed excitable systems.
For two-dimensional models, excitability can be discussed in phase space in a transparent manner. We will pose the following questions. What are the conditions for a threshold behavior? If there is no sharp threshold, what are the conditions for a regime of high (threshold-like) sensitivity? We will see that type I models indeed have a threshold whereas type II models have not. On the other hand, even type II models can show threshold-like behavior if the dynamics of w is considerably slower than that of u.
Throughout this section we use the following stimulation paradigm. We assume
that the neuron is at rest (or in a known state) and apply a short current
pulse
I(t) = q ![]() (t) of amplitude q > 0. The input pulse
influences the neuronal dynamics via Eq. (3.2). As a consequence,
the voltage u jumps at t = 0 by an amount
(t) of amplitude q > 0. The input pulse
influences the neuronal dynamics via Eq. (3.2). As a consequence,
the voltage u jumps at t = 0 by an amount
![]() u = q R/
u = q R/![]() ; the time
course of the recovery variable w, on the other hand, is continuous. In the
phase plane, the current pulse therefore shifts the state (u, w) of the
system horizontally to a new value
(u +
; the time
course of the recovery variable w, on the other hand, is continuous. In the
phase plane, the current pulse therefore shifts the state (u, w) of the
system horizontally to a new value
(u + ![]() u, w). How does the system
return to equilibrium? How does the behavior depend on the amplitude q of
the current pulse?
u, w). How does the system
return to equilibrium? How does the behavior depend on the amplitude q of
the current pulse?
We will see that the behavior can depend on q in two qualitatively distinct ways. In type I models, the response to the input shows an `all-or-nothing' behavior and consists either of a significant pulse (that is, an action potential) or a simple decay back to rest. In this sense, type I models exhibit a threshold behavior. If the action potential occurs, it has always roughly the same amplitude, but occurs at different delays depending on the strength q of the stimulating current pulse. In type II models, on the other hand, the amplitude of the response depends continuously on the amplitude q. Therefore, type II models do not have a sharp threshold.
Note that even in a model with threshold, a first input pulse that lifts the state of the system above the threshold can be counterbalanced by a second negative input which pulls the state of the system back. Thus, even in models with a threshold, the threshold is only `seen' for the specific input scenario considered here, viz., one isolated short current pulse.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
...
\par\includegraphics[width=\textwidth]{phase_type_I_detail.eps}
\end{minipage}](img339.gif) |
As discussed above, type I models are characterized by a set of three fixed
points, a stable one to the right, a saddle point in the middle, and an
unstable one to the left. The linear stability analysis at the saddle point
reveals, by definition of a saddle, one positive and one negative eigenvalue,
![]() and
and ![]() , respectively. The imaginary part of the
eigenvalues vanishes. Associated with
, respectively. The imaginary part of the
eigenvalues vanishes. Associated with ![]() is the (real) eigenvector
is the (real) eigenvector
![]() . A trajectory which approaches the saddle in direction of
. A trajectory which approaches the saddle in direction of
![]() from either side will eventually converge towards the fixed point.
There are two of these trajectories. The first one starts at infinity and
approaches the saddle from below. The second one starts at the unstable fixed
point and approaches the saddle from above. The two together define the
stable manifold of the fixed point (Verhulst, 1996; Hale and Koçak, 1991). A perturbation
around the fixed point that lies on the stable manifold returns to the fixed
point. All other perturbations will grow exponentially.
from either side will eventually converge towards the fixed point.
There are two of these trajectories. The first one starts at infinity and
approaches the saddle from below. The second one starts at the unstable fixed
point and approaches the saddle from above. The two together define the
stable manifold of the fixed point (Verhulst, 1996; Hale and Koçak, 1991). A perturbation
around the fixed point that lies on the stable manifold returns to the fixed
point. All other perturbations will grow exponentially.
The stable manifold plays an important role for the excitability of the system. Due to the uniqueness of solutions of differential equations, trajectories cannot cross. This implies that all trajectories with initial conditions to the right of the stable manifold must make a detour around the unstable fixed point before they can reach the stable fixed point. Trajectories with initial conditions to the left of the stable manifold return immediately towards the stable fixed point; cf. Fig. 3.9.
Let us now apply these considerations to models of neurons. At rest, the neuron model is at the stable fixed point. A short input current pulse moves the state of the system to the right. If the current pulse is small, the new state of the system is to the left of the stable manifold. Hence the membrane potential u decays back to rest. If the current pulse is sufficiently strong, it will shift the state of the system to the right of the stable manifold. Since the resting point is the only stable fixed point, the neuron model will eventually return to the resting potential. To do so, it has, however, to take a large detour which is seen as a pulse in the voltage variable u. The stable manifold thus acts as a threshold for spike initiation.
For I < 0 on the right-hand side of
Eq. (3.30), the phase equation
d![]() /dt has two fixed points. The
resting state
is at the stable fixed point
/dt has two fixed points. The
resting state
is at the stable fixed point
![]() =
= ![]() . The unstable fixed point at
. The unstable fixed point at
![]() =
= ![]() acts as a threshold;
cf. Fig. 3.8.
acts as a threshold;
cf. Fig. 3.8.
Let us now assume initial conditions slightly above threshold, viz.,
![]() =
= ![]() +
+ ![]()
![]() . Since
d
. Since
d![]() /dt|
/dt|![]() > 0 the system
starts to fire an action potential but for
> 0 the system
starts to fire an action potential but for
![]()
![]()
![]() 1 the phase
velocity is still close to zero and the maximum of the spike (corresponding to
1 the phase
velocity is still close to zero and the maximum of the spike (corresponding to
![]() =
= ![]() ) is reached only after a long delay. This delay depends critically
on the initial condition.
) is reached only after a long delay. This delay depends critically
on the initial condition.
In contrast to type I models, Type II models do not have a stable manifold and, hence, there is no `forbidden line' that acts as a sharp threshold. Instead of the typical all-or-nothing behavior of type I models there is a continuum of trajectories; see Fig. 3.10A.
Nevertheless, if the time scale of the u dynamics is much faster than that of the w-dynamics, then there is a critical regime where the sensitivity to the amplitude of the input current pulse can be extremely high. If the amplitude of the input pulse is increased by a tiny amount, the amplitude of the response increases a lot (`soft' threshold).
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
...dth=\textwidth]{type_II_treshold_b.eps}
% \{pout.dat.FN-9b.eps\}
\end{minipage}](img344.gif) |
In practice, the consequences of a sharp and a `soft' threshold are similar. There is, however, a subtle difference in the timing of the response between type I and type II models. In type II models, the peak of the response is always reached with roughly the same delay, independently of the size of the input pulse. It is the amplitude of the response that increases rapidly but continuously; see Fig. 3.10B. On the other hand, in type I model the amplitude of the response is rather stereotyped: either there is an action potential or not. For input currents which are just above threshold, the action potential occurs, however, with an extremely long delay. The long delay is due to the fact that the trajectory starts in the region where the two fixed points (saddle and node) have just disappeared, i.e., in a region where the velocity in phase space is very low.
Consider the generic two-dimensional neuron model given by Eqs.
(3.2) and (3.3). We measure time in units of ![]() and
take R = 1. Equations (3.2) and (3.3) are then
and
take R = 1. Equations (3.2) and (3.3) are then
| = | F(u, w) + I | (3.31) | |
| = | (3.32) |
What are the consequences of the large difference of time scales for the phase
portrait of the system? Recall that the flow is in direction of
(![]() ,
,![]() ). In the limit of
). In the limit of
![]()
![]() 0, all arrows in the flow
field are therefore horizontal, except those in the neighborhood of the
u-nullcline. On the u-nullcline,
0, all arrows in the flow
field are therefore horizontal, except those in the neighborhood of the
u-nullcline. On the u-nullcline, ![]() = 0 and arrows are vertical as
usual. Their length, however, is only of order
= 0 and arrows are vertical as
usual. Their length, however, is only of order ![]() . Intuitively
speaking, the horizontal arrows rapidly push the trajectory towards the
u-nullcline. Only close to the u-nullcline directions of movement other
than horizontal are possible. Therefore, trajectories slowly follow the
u-nullcline, except at the knees of the nullcline where they jump to a
different branch.
. Intuitively
speaking, the horizontal arrows rapidly push the trajectory towards the
u-nullcline. Only close to the u-nullcline directions of movement other
than horizontal are possible. Therefore, trajectories slowly follow the
u-nullcline, except at the knees of the nullcline where they jump to a
different branch.
![\includegraphics[width=55mm]{phase-8.eps}](img346.gif) |
Excitability can now be discussed with the help of Fig. 3.11. A
current pulse shifts the state of the system horizontally away from the stable
fixed point. If the current pulse is small, the system returns immediately
(i.e., on the fast time scale) to the stable fixed point. If the current
pulse is large enough so as to put the system beyond the middle branch of the
u-nullcline, then the trajectory is pushed towards the right branch of the
u nullcline. The trajectory follows the u-nullcline slowly upwards until
it jumps back (on the fast time scale) to the left branch of the
u-nullcline. The `jump' between the branches of the nullcline correspond to
a rapid voltage change. In terms of neuronal modeling, the jump from the
right to the left branch corresponds to downstroke of the action potential.
The middle branch of the u nullcline (where ![]() > 0) acts as a threshold
for spike initiation. This is shown in the simulation of the FitzHugh-Nagumo
model in Fig. 3.12.
> 0) acts as a threshold
for spike initiation. This is shown in the simulation of the FitzHugh-Nagumo
model in Fig. 3.12.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=60mm]{pout....
...}
{\bf B}
\par\includegraphics[width=60mm]{pout.dat.FN-12b.eps}
\end{minipage}](img347.gif) |
Let us study the piecewise linear model shown in Fig. 3.13,
The rest state is at u = w = 0. Suppose that the system is stimulated by a
short current pulse that shifts the state of the system horizontally. As long
as u < 1, we have f (u) < 0. According to (3.33), ![]() < 0 and
u returns to the rest state. For u < 0.5 the relaxation to rest is
exponential with
u(t) = exp(a t) in the limit of
< 0 and
u returns to the rest state. For u < 0.5 the relaxation to rest is
exponential with
u(t) = exp(a t) in the limit of
![]()
![]() 0. Thus,
the return to rest after a small perturbation is governed by the fast
time scale.
0. Thus,
the return to rest after a small perturbation is governed by the fast
time scale.
If the current pulse moves u to a value larger than unity, we have
![]() = f (u) > 0. Hence the voltage u increases and a pulse is emitted. That is to
say, u = 1 acts as a threshold.
= f (u) > 0. Hence the voltage u increases and a pulse is emitted. That is to
say, u = 1 acts as a threshold.
Let us now assume that an input spike from a presynaptic neuron j arrives at
time tj(f). Spike reception at the neuron generates a sub-threshold current
pulse
I(t) = c ![]() (t - tj(f)), where 0 < c
(t - tj(f)), where 0 < c ![]() 1 is the amplitude of the
pulse. The perturbation causes a voltage response,
1 is the amplitude of the
pulse. The perturbation causes a voltage response,
![\includegraphics[width=55mm]{phase-11.eps}](img352.gif) |
We have argued above that, during the pulse, the trajectory is always pushed
towards the u-nullcline. We will show in this paragraph that the trajectory
(u(t), w(t)) of the piecewise linear system
(3.33)-(3.34) follows the u-nullcline w = f (u) + I
at a distance of order ![]() . We set
. We set
To do so we use Eq. (3.36) on the right-hand-side of
(3.33) and (3.34). Thus, along the trajectory
Let us study the relaxation to the stable fixed point after a pulse in the piecewise linear model. We use f' = a for the slope of the u-nullcline and b for the slope of the w-nullcline. Evaluation of (3.40) gives
Similarly, the voltage during the spike is given by integrating
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
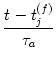
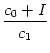 for u > 1.5 .
for u > 1.5 .