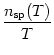The mammalian brain contains more than 1010 densely packed neurons that are conected to an intricate network. In every small volume of cortex, thousands of spikes are emitted each millisecond. An example of a spike train recording from thirty neurons is shown in Fig. 1.8. What is the information contained in such a spatio-temporal pattern of pulses? What is the code used by the neurons to transmit that information? How might other neurons decode the signal? As external observers, can we read the code and understand the message of the neuronal activity pattern?
![\centerline{
\includegraphics[width=80mm]{Figs-ch1-intro/krueger.new.eps}}](img42.gif)
|
The above questions point to the problem of neuronal coding, one of the fundamental issues in neuroscience. At present, a definite answer to these questions is not known. Traditionally it has been thought that most, if not all, of the relevant information was contained in the mean firing rate of the neuron. The firing rate is usually defined by a temporal average; see Fig. 1.9. The experimentalist sets a time window of, say T = 100ms or T = 500ms and counts the number of spikes nsp(T) that occur in this time window. Division by the length of the time window gives the mean firing rate
The concept of mean firing rates has been successfully applied during the last 80 years. It dates back to the pioneering work of Adrian (Adrian, 1928,1926) who showed that the firing rate of stretch receptor neurons in the muscles is related to the force applied to the muscle. In the following decades, measurement of firing rates became a standard tool for describing the properties of all types of sensory or cortical neurons (Mountcastle, 1957; Hubel and Wiesel, 1959), partly due to the relative ease of measuring rates experimentally. It is clear, however, that an approach based on a temporal average neglects all the information possibly contained in the exact timing of the spikes. It is therefore no surprise that the firing rate concept has been repeatedly criticized and is subject of an ongoing debate (Oram et al., 1999; Abeles, 1994; Hopfield, 1995; Bialek et al., 1991; Shadlen and Newsome, 1994; Softky, 1995; Rieke et al., 1996).
During recent years, more and more experimental evidence has accumulated which suggests that a straightforward firing rate concept based on temporal averaging may be too simplistic to describe brain activity. One of the main arguments is that reaction times in behavioral experiments are often too short to allow long temporal averages. Humans can recognize and respond to visual scenes in less than 400ms (Thorpe et al., 1996). Recognition and reaction involve several processing steps from the retinal input to the finger movement at the output. If, at each processing step, neurons had to wait and perform a temporal average in order to read the message of the presynaptic neurons, the reaction time would be much longer.
In experiments on a visual neuron in the fly, it was possible to `read the neural code' and reconstruct the time-dependent stimulus based on the neuron's firing times (Bialek et al., 1991). There is evidence of precise temporal correlations between pulses of different neurons (Abeles, 1994; Lestienne, 1996) and stimulus dependent synchronization of the activity in populations of neurons (Eckhorn et al., 1988; Singer, 1994; Gray and Singer, 1989; Gray et al., 1989; Engel et al., 1991a). Most of these data are inconsistent with a naïve concept of coding by mean firing rates where the exact timing of spikes should play no role.
In the following sections, we review some potential coding schemes and ask: What exactly is a pulse code - and what is a rate code? The question of neuronal coding has important implications for modeling, because pulse codes require a more detailed description of neuronal dynamics than rate codes. Models of neurons at different levels of detail will be the topic of part I of the book.
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.