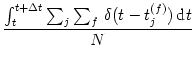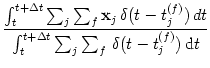A quick glance at the experimental literature reveals that there is no unique and well-defined concept of `mean firing rate'. In fact, there are at least three different notions of rate which are often confused and used simultaneously. The three definitions refer to three different averaging procedures: either an average over time, or an average over several repetitions of the experiment, or an average over a population of neurons. The following three subsections will reconsider the three concepts. An excellent discussion of rate codes can be found in (Rieke et al., 1996).
The first and most commonly used definition of a firing rate refers to a temporal average. As discussed in the preceding section, this is essentially the spike count in an interval of duration T divided by T; see Fig. 1.9. The length T of the time window is set by the experimenter and depends on the type of neuron recorded from and the stimulus. In practice, to get sensible averages, several spikes should occur within the time window. Typical values are T = 100 ms or T = 500 ms, but the duration may also be longer or shorter.
![\hbox{{\bf A} \hspace{75mm} {\bf B}}
\hbox{
\includegraphics[width=75mm]{Figs-ch...
...ps}
\hspace{10mm}
\includegraphics[width=50mm]{Figs-ch1-intro/rate-gainf.eps}
}](img45.gif) |
This definition of rate has been successfully used in many preparations, particularly in experiments on sensory or motor systems. A classical example is the stretch receptor in a muscle spindle (Adrian, 1926). The number of spikes emitted by the receptor neuron increases with the force applied to the muscle. Another textbook example is the touch receptor in the leech (Kandel and Schwartz, 1991). The stronger the touch stimulus, the more spikes occur during a stimulation period of 500 ms.
These classical results show that the experimenter as an external observer can evaluate and classify neuronal firing by a spike count measure - but is this really the code used by neurons in the brain? In other words, is a neuron which receives signals from a sensory neuron only looking at and reacting to the number of spikes it receives in a time window of, say, 500 ms? We will approach this question from a modeling point of view later on in the book. Here we discuss some critical experimental evidence.
From behavioral experiments it is known that reaction times are often rather short. A fly can react to new stimuli and change the direction of flight within 30-40 ms; see the discussion in (Rieke et al., 1996). This is not long enough for counting spikes and averaging over some long time window. The fly has to respond after a postsynaptic neuron has received one or two spikes. Humans can recognize visual scenes in just a few hundred milliseconds (Thorpe et al., 1996), even though recognition is believed to involve several processing steps. Again, this does not leave enough time to perform temporal averages on each level. In fact, humans can detect images in a sequence of unrelated pictures even if each image is shown for only 14 - 100 milliseconds (Keysers et al., 2001).
Temporal averaging can work well in cases where the stimulus is constant or slowly varying and does not require a fast reaction of the organism - and this is the situation usually encountered in experimental protocols. Real-world input, however, is hardly stationary, but often changing on a fast time scale. For example, even when viewing a static image, humans perform saccades, rapid changes of the direction of gaze. The image projected onto the retinal photo receptors changes therefore every few hundred milliseconds.
Despite its shortcomings, the concept of a firing rate
code is widely used not only in experiments,
but also in models of neural networks.
It has led to the idea that a neuron transforms information
about a single input variable
(the stimulus strength)
into a single continuous output variable (the firing rate);
cf. Fig. 1.9B.
The output rate ![]() increases with the stimulus
strength and saturates for large input I0
towards a maximum value
increases with the stimulus
strength and saturates for large input I0
towards a maximum value
![]() .
In experiments, a single neuron can be stimulated
by injecting with an intra-cellular electrode
a constant current I0.
The relation between the measured firing frequency
.
In experiments, a single neuron can be stimulated
by injecting with an intra-cellular electrode
a constant current I0.
The relation between the measured firing frequency ![]() and the applied input current I0
is sometimes called the frequency-current curve of
the neuron. In models, we formalize
the relation between firing frequency (rate)
and input current and write
and the applied input current I0
is sometimes called the frequency-current curve of
the neuron. In models, we formalize
the relation between firing frequency (rate)
and input current and write
![]() = g(I0). We
refer to g as the neuronal gain
function or transfer function.
= g(I0). We
refer to g as the neuronal gain
function or transfer function.
From the point of view of rate coding, spikes are just a convenient way
to transmit the analog output variable ![]() over long distances.
In fact, the best coding scheme to transmit the
value of the rate
over long distances.
In fact, the best coding scheme to transmit the
value of the rate ![]() would be by a regular
spike train with intervals 1/
would be by a regular
spike train with intervals 1/![]() .
In this case, the rate could be reliably measured
after only two spikes. From the point of view of rate
coding, the irregularities
encountered in real spike trains of neurons in the cortex
must therefore be considered as noise.
In order to get rid of the noise and arrive at
a reliable estimate of the rate,
the experimenter (or the postsynaptic neuron)
has to average over a larger number of spikes.
A critical discussion of the temporal averaging concept can be found
in
(Shadlen and Newsome, 1994; Softky, 1995; Rieke et al., 1996).
.
In this case, the rate could be reliably measured
after only two spikes. From the point of view of rate
coding, the irregularities
encountered in real spike trains of neurons in the cortex
must therefore be considered as noise.
In order to get rid of the noise and arrive at
a reliable estimate of the rate,
the experimenter (or the postsynaptic neuron)
has to average over a larger number of spikes.
A critical discussion of the temporal averaging concept can be found
in
(Shadlen and Newsome, 1994; Softky, 1995; Rieke et al., 1996).
There is a second definition of rate which works for stationary
as well as for time-dependent stimuli.
The experimenter records from a neuron
while stimulating with some input sequence.
The same stimulation sequence
is repeated several times and the neuronal response
is reported in a Peri-Stimulus-Time Histogram (PSTH); see Fig. 1.10.
The time t is measured with respect to the
start of the stimulation sequence and
![]() t is typically in the range of one or
a few milliseconds.
The number of occurrences of spikes
nK(t;t +
t is typically in the range of one or
a few milliseconds.
The number of occurrences of spikes
nK(t;t + ![]() t)
summed over all repetitions of the experiment divided by
the number K of repetitions is a measure
of the typical activity of the neuron
between time t and
t +
t)
summed over all repetitions of the experiment divided by
the number K of repetitions is a measure
of the typical activity of the neuron
between time t and
t + ![]() t.
A further division by the interval length
t.
A further division by the interval length ![]() t
yields the spike density of the PSTH
t
yields the spike density of the PSTH
![\centerline{
\includegraphics[width=100mm]{Figs-ch1-intro/rate-b.eps}}](img51.gif)
|
As an experimental procedure, the spike density measure is a useful method to evaluate neuronal activity, in particular in the case of time-dependent stimuli. The obvious problem with this approach is that it can not be the decoding scheme used by neurons in the brain. Consider for example a frog which wants to catch a fly. It can not wait for the insect to fly repeatedly along exactly the same trajectory. The frog has to base its decision on a single `run' - each fly and each trajectory is different.
Nevertheless, the experimental spike density measure can make sense, if there are large populations of independent neurons that receive the same stimulus. Instead of recording from a population of N neurons in a single run, it is experimentally easier to record from a single neuron and average over N repeated runs. Thus, the spike density coding relies on the implicit assumption that there are always populations of neurons and therefore leads us to the third notion of a firing rate, viz., a rate defined as a population average.
![\hbox{{\bf A} \hspace{55mm} {\bf B}}
\hbox{\hspace{5mm}
\includegraphics[width=5...
...e-d.eps}
\hspace{5mm}
\includegraphics[width=70mm]{Figs-ch1-intro/rate-c.eps}
}](img52.gif)
|
The number of neurons in the brain is huge. Often many neurons have similar properties and respond to the same stimuli. For example, neurons in the primary visual cortex of cats and monkeys are arranged in columns of cells with similar properties (Hubel, 1988; Hubel and Wiesel, 1962,1977). Let us idealize the situation and consider a population of neurons with identical properties. In particular, all neurons in the population should have the same pattern of input and output connections. The spikes of the neurons in a population m are sent off to another population n. In our idealized picture, each neuron in population n receives input from all neurons in population m. The relevant quantity, from the point of view of the receiving neuron, is the proportion of active neurons in the presynaptic population m; see Fig. 1.11A. Formally, we define the population activity
The population activity may vary rapidly and can reflect changes in the stimulus conditions nearly instantaneously (Brunel et al., 2001; Gerstner, 2000a). Thus the population activity does not suffer from the disadvantages of a firing rate defined by temporal averaging at the single-unit level. A potential problem with the definition (1.10) is that we have formally required a homogeneous population of neurons with identical connections which is hardly realistic. Real populations will always have a certain degree of heterogeneity both in their internal parameters and in their connectivity pattern. Nevertheless, rate as a population activity (of suitably defined pools of neurons) may be a useful coding principle in many areas of the brain. For inhomogeneous populations, the definition (1.10) may be replaced by a weighted average over the population.
We give an example of a weighted average in an
inhomogeneous population.
Let us suppose that we are studying a population of neurons
which respond to a stimulus ![]() .
We may think of
.
We may think of ![]() as the location of the stimulus
in input space.
Neuron i responds best to stimulus
as the location of the stimulus
in input space.
Neuron i responds best to stimulus ![]() ,
another neuron j responds best to stimulus
,
another neuron j responds best to stimulus ![]() .
In other words, we may say
that the spikes for
a neuron i `represent' an input vector
.
In other words, we may say
that the spikes for
a neuron i `represent' an input vector ![]() and those of j an input vector
and those of j an input vector ![]() .
In a large population, many neurons
will be active simultaneously when
a new stimulus
.
In a large population, many neurons
will be active simultaneously when
a new stimulus ![]() is represented.
The location of this stimulus
can then be estimated from the
weighted population average
is represented.
The location of this stimulus
can then be estimated from the
weighted population average
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
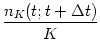 .
.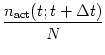 =
=