




Next: 6.2 Density Equations
Up: 6. Population Equations
Previous: 6. Population Equations
6.1 Fully Connected Homogeneous Network
Figure 6.1:
Population of neurons (schematic). All neurons
receive the same input
Iext(t) (left)
which results in a time dependent
population activity A(t) (right).
![\hbox{\hspace{20mm}
\includegraphics[width=80mm]{Figs-ch-pop/pop-1.eps}}](img966.gif) |
We study a large and homogeneous population of neurons; cf. Fig. 6.1. By homogeneous we mean that all neurons
1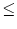 iN
are identical and receive the same external input
Iiext(t) = Iext(t). Moreover, in a homogeneous population, the interaction
strength between the neurons is taken to be uniform,
iN
are identical and receive the same external input
Iiext(t) = Iext(t). Moreover, in a homogeneous population, the interaction
strength between the neurons is taken to be uniform,
wij = 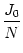 , , |
(6.2) |
where J0 is a parameter. For J0 = 0 all neurons are independent; a
value J0 > 0 (J0 < 0) implies excitatory (inhibitory) all-to-all coupling.
The interaction strength scales with one over the number of neurons so that
the total synaptic input to each neuron remains finite in the limit of
N
 .
.
Model neurons are described by formal spiking neurons as introduced in
Chapter 4. In the case of leaky integrate-and-fire neurons with
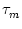 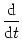 ui = - ui + R Ii(t) ui = - ui + R Ii(t) |
(6.3) |
a homogeneous network implies that all neurons have the same input resistance
R, the same membrane time constant 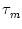 , as well as identical threshold
and reset values. The input current Ii takes care of both the external
drive and synaptic coupling
, as well as identical threshold
and reset values. The input current Ii takes care of both the external
drive and synaptic coupling
Ii = 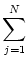 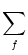 wij wij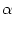 (t - tj(f)) + Iext(t) . (t - tj(f)) + Iext(t) . |
(6.4) |
Here we have assumed that each input spike generates a postsynaptic current
with some generic time course
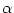 (t - tj(f)). The sum on the right-hand
side of (6.4) runs over all firing times of all neurons.
Because of the homogeneous all-to-all coupling, the total input current is
identical for all neurons. To see this, we insert Eq. (6.2) and use
the definition of the population activity, Eq. (6.1). We find a
total input current,
(t - tj(f)). The sum on the right-hand
side of (6.4) runs over all firing times of all neurons.
Because of the homogeneous all-to-all coupling, the total input current is
identical for all neurons. To see this, we insert Eq. (6.2) and use
the definition of the population activity, Eq. (6.1). We find a
total input current,
I(t) = J0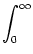 (s) A(t - s) ds + Iext(t) , (s) A(t - s) ds + Iext(t) , |
(6.5) |
which is independent of the neuronal index i. As an aside we note that for
conductance-based synaptic input, the total input current would depend on the
neuronal membrane potential which is different from one neuron to the next.
Instead of the integrate-and-fire neuron, we may also use the Spike Response
Model (SRM) as the elementary unit of the population. The membrane potential
of a SRM neuron is of the form
ui(t) = 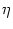 (t - (t - 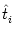 ) + hPSP(t|) , ) + hPSP(t|) , |
(6.6) |
where 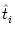 is the most recent firing time of neuron i. The kernel
is the most recent firing time of neuron i. The kernel
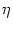 (.) describes the spike and the spike after-potential while
(.) describes the spike and the spike after-potential while
hPSP(t|) = 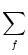 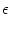 (t - , t - tj(f)) + (t - , t - tj(f)) + 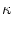 (t - , s) Iext(t - s) ds (t - , s) Iext(t - s) ds |
(6.7) |
is the postsynaptic potential caused by firings tj(f) of presynaptic neurons
j or by external input
Iext(t). The kernel 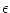 models the
response of the neuron to a single presynaptic input spike while
models the
response of the neuron to a single presynaptic input spike while 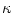 is the response to a unit current pulse. In Eq. (6.7) we have already
exploited Eq. (6.2) and replaced the synaptic efficacies wij by
J0/N. The population theory developed in this chapter is valid for
arbitrary response kernels and and for a broad variety
of refractory kernels . By an appropriate choice of the kernels, we
recover the integrate-and-fire model; cf. Chapter 4.2. If we
suppress the
t - dependency of and , we recover
the simple model SRM0 from Chapter 4.2.3.
is the response to a unit current pulse. In Eq. (6.7) we have already
exploited Eq. (6.2) and replaced the synaptic efficacies wij by
J0/N. The population theory developed in this chapter is valid for
arbitrary response kernels and and for a broad variety
of refractory kernels . By an appropriate choice of the kernels, we
recover the integrate-and-fire model; cf. Chapter 4.2. If we
suppress the
t - dependency of and , we recover
the simple model SRM0 from Chapter 4.2.3.
Similarly to the approach that we used for the total input current of an
integrate-and-fire neuron, we can rewrite Eq. (6.7) in terms of the
population activity A,
hPSP(t|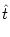 ) = J0(t - , s) A(t - s) ds + (t - , s) Iext(t - s) ds . ) = J0(t - , s) A(t - s) ds + (t - , s) Iext(t - s) ds . |
(6.8) |
Thus, given the activity A(t') and the external input
Iext for
t' < t we can determine the potential
hPSP(t|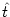 ) of a neuron that
has fired its last spike at . Note that we have suppressed the index
i, since all neurons that have fired their last spike at have the
same postsynaptic potential
hPSP. As above, this is an
immediate consequence of the assumption of a homogeneous network and does not
require a limit of
N.
) of a neuron that
has fired its last spike at . Note that we have suppressed the index
i, since all neurons that have fired their last spike at have the
same postsynaptic potential
hPSP. As above, this is an
immediate consequence of the assumption of a homogeneous network and does not
require a limit of
N.
In the absence of noise, the next firing time of a spiking neuron i is found
from the threshold condition,
ui(t) = 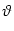 and ui > 0 . and ui > 0 . |
(6.9) |
In the presence of noise, the next firing time of a given neuron i cannot be
predicted in a deterministic fashion. In the case of integrate-and-fire
neurons with diffusive noise (stochastic spike arrival), a large noise level
leads to a broad distribution of the membrane potential and indirectly to a
large distribution of interspike intervals; cf. Chapter 5. In
the case of spiking neurons with escape noise (noisy threshold), firing occurs
probabilistically which results in a similar large distribution of interspike
intervals. In the following sections, we formulate population equations for
various types of spiking neuron and different types of noise. We start in the
next section with a population of integrate-and-fire neurons with diffusive
noise and turn then to Spike Response Model neurons.
Next: 6.2 Density Equations
Up: 6. Population Equations
Previous: 6. Population Equations
Gerstner and Kistler
Spiking Neuron Models. Single Neurons, Populations, Plasticity
Cambridge University Press, 2002
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.