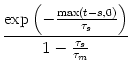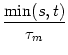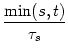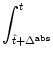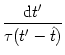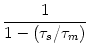The Spike Response Model (SRM) is - just like the nonlinear integrate-and-fire model - a generalization of the leaky integrate-and-fire model. The direction of the generalization is, however, somewhat different. In the nonlinear integrate-and-fire model, parameters are made voltage dependent whereas in the SRM they depend on the time since the last output spike. Another difference between integrate-and-fire models and the SRM concerns the formulation of the equations. While integrate-and-fire models are usually defined in terms of differential equations, the SRM expresses the membrane potential at time t as an integral over the past.
The explicit dependence of the membrane potential upon the last output spike allows us to model refractoriness as a combination of three components, viz., (i) a reduced responsiveness after an output spike; (ii) an increase in threshold after firing; and (iii) a hyperpolarizing spike after-potential. In Section 4.2.1 the Spike Response Model is introduced and its properties illustrated. Its relation to the integrate-and-fire model is the topic of Section 4.2.2. An important special case of the Spike Response Model is the simplified model SRM0 that we have already encountered in Chapter 1.3.1. Section 4.2.3 will discuss it in more detail.
In the framework of the Spike Response Model
the state of a neuron i is described by a single variable ui. In the
absence of spikes, the variable ui is at its resting value,
urest = 0. Each incoming spike will perturb ui and
it takes some time before ui returns to zero. The function ![]() describes the time course of the response to an incoming spike. If, after the
summation of the effects of several incoming spikes, ui reaches the
threshold
describes the time course of the response to an incoming spike. If, after the
summation of the effects of several incoming spikes, ui reaches the
threshold ![]() an output spike is triggered. The form of the action
potential and the after-potential is described by
a function
an output spike is triggered. The form of the action
potential and the after-potential is described by
a function ![]() . Let us suppose neuron i has fired its last spike at time
. Let us suppose neuron i has fired its last spike at time
![]() . After firing the evolution of ui is given by
. After firing the evolution of ui is given by
In contrast to the integrate-and-fire neuron discussed in
Section (4.1) the threshold ![]() is not fixed but may also
depend on
t -
is not fixed but may also
depend on
t - ![]()
| (4.25) |
| (4.27) |
So far Eqs. (4.1) and (4.24) define a mathematical model. Can we
give a biological interpretation of the terms? Let us identify the variable
ui with the membrane potential of neuron i. The functions ![]() ,
,
![]() and
and
![]() are response kernels that describe
the effect of spike emission and spike reception on the variable ui. This
interpretation has motivated the name `Spike Response Model', SRM for short
(Gerstner, 1995; Kistler et al., 1997). Let us discuss the meaning of the response
kernels.
are response kernels that describe
the effect of spike emission and spike reception on the variable ui. This
interpretation has motivated the name `Spike Response Model', SRM for short
(Gerstner, 1995; Kistler et al., 1997). Let us discuss the meaning of the response
kernels.
![\centerline{\includegraphics[width=60mm]{SRM-schema.eps}}](img448.gif) |
The kernel ![]() describes the standard form of an action potential of
neuron i including the negative overshoot which typically follows a spike
(after-potential). Graphically speaking, a contribution
describes the standard form of an action potential of
neuron i including the negative overshoot which typically follows a spike
(after-potential). Graphically speaking, a contribution ![]() is `pasted
in' each time the membrane potential reaches the threshold
is `pasted
in' each time the membrane potential reaches the threshold ![]() ; cf.
Fig. 4.5. Since the form of the spike is always the same, the exact
time course of the action potential carries no information. What matters is
whether there is the event `spike' or not. The event is fully characterized
by the firing time ti(f). In a simplified model, the form of the
action potential may therefore be neglected as long as we keep track of the
firing times ti(f). The kernel
; cf.
Fig. 4.5. Since the form of the spike is always the same, the exact
time course of the action potential carries no information. What matters is
whether there is the event `spike' or not. The event is fully characterized
by the firing time ti(f). In a simplified model, the form of the
action potential may therefore be neglected as long as we keep track of the
firing times ti(f). The kernel ![]() describes then simply the `reset' of
the membrane potential to a lower value after the spike at
describes then simply the `reset' of
the membrane potential to a lower value after the spike at ![]() just like
in the integrate-and-fire model. The leaky integrate-and-fire model is in
fact a special case of the SRM as we will see below in
Section 4.2.2.
just like
in the integrate-and-fire model. The leaky integrate-and-fire model is in
fact a special case of the SRM as we will see below in
Section 4.2.2.
The kernel
![]() (t -
(t - ![]() , s) is the linear response of the
membrane potential to an input current. It describes the time course of a
deviation of the membrane potential from its resting value that is caused by a
short current pulse (``impulse response''). We have
already seen in Chapters 2.2 and 3 that the response
depends, in general, on the time that has passed since the last output spike
at
, s) is the linear response of the
membrane potential to an input current. It describes the time course of a
deviation of the membrane potential from its resting value that is caused by a
short current pulse (``impulse response''). We have
already seen in Chapters 2.2 and 3 that the response
depends, in general, on the time that has passed since the last output spike
at ![]() . Immediately after
. Immediately after ![]() many ion channels are open so that
the resistance of the membrane is reduced. The voltage response to an input
current pulse decays therefore more rapidly back to zero than in a neuron that
has been inactive. A reduced or shorter response is one of the signatures of
neuronal refractoriness. This form of refractory effect is taken care of by
making the kernel
many ion channels are open so that
the resistance of the membrane is reduced. The voltage response to an input
current pulse decays therefore more rapidly back to zero than in a neuron that
has been inactive. A reduced or shorter response is one of the signatures of
neuronal refractoriness. This form of refractory effect is taken care of by
making the kernel ![]() depend, via its first argument, on the time
difference
t -
depend, via its first argument, on the time
difference
t - ![]() . We illustrate the idea in Fig. 4.5. The
response to a first input pulse at t' is shorter and less pronounced than
that to a second one at t'', an effect which is well-known experimentally
(Fuortes and Mantegazzini, 1962; Stevens and Zador, 1998; Powers and Binder, 1996).
. We illustrate the idea in Fig. 4.5. The
response to a first input pulse at t' is shorter and less pronounced than
that to a second one at t'', an effect which is well-known experimentally
(Fuortes and Mantegazzini, 1962; Stevens and Zador, 1998; Powers and Binder, 1996).
The kernel
![]() (t -
(t - ![]() , s) as a function of
s = t - tj(f) can be
interpreted as the time course of a postsynaptic potential
evoked by the firing of a presynaptic neuron
j at time tj(f). Depending on the sign of the synapse from j, to i,
, s) as a function of
s = t - tj(f) can be
interpreted as the time course of a postsynaptic potential
evoked by the firing of a presynaptic neuron
j at time tj(f). Depending on the sign of the synapse from j, to i,
![]() models either an excitatory or inhibitory postsynaptic
potential (EPSP or IPSP). Similarly as for the kernel
models either an excitatory or inhibitory postsynaptic
potential (EPSP or IPSP). Similarly as for the kernel ![]() , the exact
shape of the postsynaptic potential depends on the time
t -
, the exact
shape of the postsynaptic potential depends on the time
t - ![]() that has
passed since the last spike of the postsynaptic neuron i. In particular, if
neuron i has been active immediately before the arrival of a presynaptic
action potential, the postsynaptic neuron is in a state of refractoriness. In
this case, the response to an input spike is smaller than that of an
`unprimed' neuron. The first argument of
that has
passed since the last spike of the postsynaptic neuron i. In particular, if
neuron i has been active immediately before the arrival of a presynaptic
action potential, the postsynaptic neuron is in a state of refractoriness. In
this case, the response to an input spike is smaller than that of an
`unprimed' neuron. The first argument of
![]() (t -
(t - ![]() , s) accounts
for the dependence upon the last firing time of the postsynaptic neuron.
, s) accounts
for the dependence upon the last firing time of the postsynaptic neuron.
In order to simplify the notation for later use, it is convenient to introduce the total postsynaptic potential,
Refractoriness may be characterized experimentally by the observation that immediately after a first action potential it is impossible (absolute refractoriness) or more difficult (relative refractoriness) to excite a second spike (Fuortes and Mantegazzini, 1962).
Absolute refractoriness can be incorporated in the SRM by setting the
dynamic threshold during a time
![]() to an extremely high value that
cannot be attained.
to an extremely high value that
cannot be attained.
Relative refractoriness can be mimicked in various ways; see
Fig. 4.5. First, after the spike the membrane potential, and hence
![]() , passes through a regime of hyperpolarization (after-potential) where
the voltage is below the resting potential. During this phase, more
stimulation than usual is needed to drive the membrane potential above
threshold. This is equivalent to a transient increase of the firing threshold
(see below). Second,
, passes through a regime of hyperpolarization (after-potential) where
the voltage is below the resting potential. During this phase, more
stimulation than usual is needed to drive the membrane potential above
threshold. This is equivalent to a transient increase of the firing threshold
(see below). Second, ![]() and
and ![]() contribute to relative
refractoriness because, immediately after an action potential, the response to
incoming spikes is shorter and, possibly, of reduced amplitude
(Fuortes and Mantegazzini, 1962). Thus more input spikes are needed to evoke the same
depolarization of the membrane potential as in an `unprimed' neuron. The
first argument of the
contribute to relative
refractoriness because, immediately after an action potential, the response to
incoming spikes is shorter and, possibly, of reduced amplitude
(Fuortes and Mantegazzini, 1962). Thus more input spikes are needed to evoke the same
depolarization of the membrane potential as in an `unprimed' neuron. The
first argument of the ![]() function (or
function (or ![]() function) allows us
to incorporate this effect.
function) allows us
to incorporate this effect.
From a formal point of view, there is no need to interpret the variable u as
the membrane potential. It is, for example, often convenient to transform the
variable u so as to remove the time-dependence of the threshold. In fact, a
general Spike Response Model with arbitrary time-dependent threshold
![]() (t -
(t - ![]() ) =
) = ![]() +
+ ![]() (t -
(t - ![]() ), can always be
transformed into a Spike Response Model with fixed threshold
), can always be
transformed into a Spike Response Model with fixed threshold
![]() by
a change of variables
by
a change of variables
In Chapter 3 we have studied the FitzHugh-Nagumo model as an
example of a two-dimensional neuron model. Here we want to show that the
response of the FitzHugh-Nagumo model to a short input current pulse depends
on the time since the last spike. Let us trigger, in a simulation of the
model, an action potential at t = 0. This can be done by applying a short,
but strong current pulse. The result is a voltage trajectory of large
amplitude which we identify with the kernel ![]() (t). Figure 4.6
shows the
hyperpolarizing spike after-potential which decays slowly back to the resting
level. To test the responsiveness of the FitzHugh-Nagumo model during the
recovery phase after the action potential, we apply, at a time t(f) > 0, a
second short input current pulse of low amplitude. The response to this test
pulse is compared with the unperturbed trajectory. The difference between the
two trajectories defines the kernel
(t). Figure 4.6
shows the
hyperpolarizing spike after-potential which decays slowly back to the resting
level. To test the responsiveness of the FitzHugh-Nagumo model during the
recovery phase after the action potential, we apply, at a time t(f) > 0, a
second short input current pulse of low amplitude. The response to this test
pulse is compared with the unperturbed trajectory. The difference between the
two trajectories defines the kernel
![]() (t -
(t - ![]() , t - t(f)). In
Fig. 4.6 several trajectories are overlayed showing the response
to stimulation at
t = 10, 15, 20, 30 or 40. The shape and duration of the
response curve depends on the time that has passed since the initiation of the
action potential. Note that the time constant of the response kernel
, t - t(f)). In
Fig. 4.6 several trajectories are overlayed showing the response
to stimulation at
t = 10, 15, 20, 30 or 40. The shape and duration of the
response curve depends on the time that has passed since the initiation of the
action potential. Note that the time constant of the response kernel
![]() is always shorter than that of the hyperpolarizing spike
after-potential. Analogous results for the Hodgkin-Huxley model will be
discussed below in Section 4.3.1.
is always shorter than that of the hyperpolarizing spike
after-potential. Analogous results for the Hodgkin-Huxley model will be
discussed below in Section 4.3.1.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=70mm]{pout....
...includegraphics[width=30mm]{pout.dat.FN-SRM-b.eps}
\end{center} \end{minipage}](img451.gif) |
![[t]
\centerline{\includegraphics[width=8cm]{Figs-ch-signal/MODULATION.eps}}](img452.gif) |
Motoneurons exhibit a rather slow return to the resting potential after an
action potential (Powers and Binder, 1996). The time constant of the
decay of the hyperpolarizing spike after-potential can be in the range of
100ms or more and is therefore much slower than the membrane time constant
that characterizes the response to a short current input. On the other hand,
it is found that if motoneurons are stimulated by a constant super-threshold
current, their membrane potential has a roughly linear trajectory when
approaching threshold. To qualitatively describe these observations, we can
use a Spike Response Model with the following kernels:
The effect of the modulation of the input conductance as a function of
t - ![]() is depicted in Fig. 4.7. An input
current pulse shortly after the reset at time
is depicted in Fig. 4.7. An input
current pulse shortly after the reset at time ![]() evokes a postsynaptic
potential of much lower amplitude than an input current pulse that arrives
much later. Fig. 4.7 qualitatively reproduces
the membrane trajectory of motoneurons when stimulated by the same input
pattern (Poliakov et al., 1996; Powers and Binder, 1996).
evokes a postsynaptic
potential of much lower amplitude than an input current pulse that arrives
much later. Fig. 4.7 qualitatively reproduces
the membrane trajectory of motoneurons when stimulated by the same input
pattern (Poliakov et al., 1996; Powers and Binder, 1996).
In this section, we show that the leaky integrate-and-fire neuron
defined in Section 4.1 is a special case of the Spike Response Model.
We consider an integrate-and-fire neuron driven by external current
Iext and postsynaptic current pulses
![]() (t - tj(f)). The potential ui
is thus given by
(t - tj(f)). The potential ui
is thus given by
| u(t) | = ur exp |
(4.34) |
| + |
||
| + |
||
| = |
In order obtain an explicit expression for the ![]() kernel
(4.36) we have to specify the time course of the postsynaptic
current
kernel
(4.36) we have to specify the time course of the postsynaptic
current ![]() (s). Here, we take
(s). Here, we take ![]() (s) as defined in (4.21),
viz.,
(s) as defined in (4.21),
viz.,
| (4.38) |
![\centerline{\includegraphics[width=6cm]{eps2_kernel.ps}}](img490.gif) |
We have seen above that the Spike Response Model contains the integrate-and-fire model as a special case. In this example, we show in addition that even a generalization of the integrate-and-fire model that has a time dependent membrane time constant can be described within the SRM framework.
To be specific, we consider an integrate-and-fire model with spike-time
dependent time constant, i.e., with a membrane time constant ![]() that
is a function of the time since the last postsynaptic spike,
that
is a function of the time since the last postsynaptic spike,
The phenomenological neuron model SRM0 introduced in Chapter 1.3.1 is a special case of the Spike Response Model. In this section we review its relation to the SRM and the integrate-and-fire model.
A simplified version of the spike response model can be constructed by
neglecting the dependence of ![]() and
and ![]() upon the first
argument. We set
upon the first
argument. We set
| = | |||
| = |
![\centerline{\includegraphics[width=80mm]{SRM1b.eps}}](img506.gif) |
The simplified model SRM0 defined in (4.42) with the ![]() kernel
defined in (4.43) can be reinterpreted as a model with a dynamic
threshold,
kernel
defined in (4.43) can be reinterpreted as a model with a dynamic
threshold,
| (4.44) |
![\centerline{
\includegraphics[width=70mm]{pout.dat.3integr.eps}} \vspace{-5mm}](img507.gif) |
The basic equation of the leaky integrate-and-fire model, Eq. (4.3), is a linear differential equation. However, because of the reset of the membrane potential after firing, the integration is not completely trivial. In fact, there are two different ways of proceeding with the integration of Eq. (4.3). In Section 4.2.2 we have treated the reset as a new initital condition and thereby constructed an exact mapping of the integrate-and-fire model to the Spike Response Model. We now turn to the second method and describe the reset as a current pulse. As we will see, the result is an approximative mapping to the simplified model SRM0.
Let us consider a short current pulse
Iiout = - q ![]() (t)
applied to the RC circuit of Fig. 4.1. It removes a charge q
from the capacitor C and lowers the potential by an amount
(t)
applied to the RC circuit of Fig. 4.1. It removes a charge q
from the capacitor C and lowers the potential by an amount
![]() u = - q/C. Thus, a reset of the membrane potential from a value of
u =
u = - q/C. Thus, a reset of the membrane potential from a value of
u = ![]() to a new value u = ur corresponds to an `output' current pulse which removes
a charge
q = C (
to a new value u = ur corresponds to an `output' current pulse which removes
a charge
q = C (![]() - ur). The reset takes place every time when
the neuron fires. The total reset current is therefore
- ur). The reset takes place every time when
the neuron fires. The total reset current is therefore
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=45mm]{pout...
...
{\bf B}
\par\includegraphics[height=45mm]{pout.dat.IF-eps.eps}
\end{minipage}](img512.gif) |
We note that, in contrast to Eq. (4.42), we still have on the right-hand
side of Eq. (4.50) a sum over past spikes of neuron i. According to
Eq. (4.51) the effect of the ![]() -kernel decays with a time
constant
-kernel decays with a time
constant ![]() . In realistic spike trains, the interval between two spikes
is typically much longer than the membrane time constant
. In realistic spike trains, the interval between two spikes
is typically much longer than the membrane time constant ![]() . Hence the
sum over the
. Hence the
sum over the ![]() terms is usually dominated by the most recent
firing time
ti(f) < t of neuron i. We therefore truncate the sum ofer f and
neglect the effect of earlier spikes,
terms is usually dominated by the most recent
firing time
ti(f) < t of neuron i. We therefore truncate the sum ofer f and
neglect the effect of earlier spikes,
A careful comparison of Eq. (4.51) with Eq. (4.35) shows that
the kernel ![]() is different from the kernel
is different from the kernel ![]() derived previously for
the exact mapping of the integrate-and-fire model to the full
Spike Response Model. The difference is most easily seen if we set the reset
potential to ur = 0. While the kernel
derived previously for
the exact mapping of the integrate-and-fire model to the full
Spike Response Model. The difference is most easily seen if we set the reset
potential to ur = 0. While the kernel ![]() in Eq. (4.35) vanishes
in this case, the kernel
in Eq. (4.35) vanishes
in this case, the kernel ![]() is nonzero. In fact, whereas in the full SRM
the reset is taken care of by the definition of
is nonzero. In fact, whereas in the full SRM
the reset is taken care of by the definition of
![]() (t -
(t - ![]() , s) and
, s) and
![]() (t -
(t - ![]() , s), the reset in the simplified model SRM0 is included
in the kernel
, s), the reset in the simplified model SRM0 is included
in the kernel ![]() . The relation between the kernels of the simplified
model SRM0 to that of the full model are discussed below in more detail.
. The relation between the kernels of the simplified
model SRM0 to that of the full model are discussed below in more detail.
If ![]() (s) is given by (4.21), then the integral on the right-hand
side of (4.52) can be done and yields
(s) is given by (4.21), then the integral on the right-hand
side of (4.52) can be done and yields
What is the relation between the ![]() kernel derived in (4.36)
and the
kernel derived in (4.36)
and the
![]() introduced in (4.52)? We will show in this
paragraph that
introduced in (4.52)? We will show in this
paragraph that
| = |
||
| = |
||
| - exp |
(4.58) |
By a completely analogous sequence of transformations it is possible to show that
Similarly we can compare the ![]() kernel in (4.35) and the
kernel in (4.35) and the
![]() kernel defined in (4.51),
kernel defined in (4.51),
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.