In this section we study the relation between detailed conductance-based neuron models and formal spiking neurons as introduced above. In Section 4.3.1, we discuss how an approximative mapping between the Spike Response Model and the Hodgkin-Huxley model can be established. While the Hodgkin-Huxley model is of type II, cortical neurons are usually described by type I models. In Subsection 4.3.2 we focus on a type-I model of cortical interneurons and reduce it systematically to different variants of spiking neuron models, in particular to a nonlinear integrate-and-fire model and a Spike Response Model. In all sections, the performance of the reduced models is compared to that of the full model. To do so we test the models with a constant or fluctuating input current.
The system of equations proposed by Hodgkin and Huxley (see Chapter 2.2) is rather complicated. It consists of four coupled nonlinear differential equations and as such is difficult to analyze mathematically. For this reason, several simplifications of the Hodgkin-Huxley equations have been proposed. The most common approach reduces the set of four differential equations to a two-dimensional problem as discussed in Chapter 3. In this section, we will take a somewhat different approach to reduce the four Hodgkin-Huxley equations to a single variable u(t), the membrane potential of the neuron (Kistler et al., 1997). As we have seen in Fig. 2.4B, the Hodgkin-Huxley model shows a sharp, threshold-like transition between an action potential (spike) for a strong stimulus and a graded response (no spike) for a slightly weaker stimulus. This suggests the idea that emission of an action potential can be described by a threshold process. We therefore aim for a reduction towards a spiking neuron model where spikes are triggered by a voltage threshold. Specifically, we will establish an approximative mapping between the Spike Response Model and the Hodgkin-Huxley model.
Action potentials in the Hodgkin-Huxley model have the stereotyped time course
shown in Fig. 2.4A. Whatever the stimulating current that has
triggered the spike, the form of the action potential is always roughly the
same (as long as the current stays in a biologically realistic regime). This
is the major observation that we will exploit in the following. Let us
consider the spike that has been triggered at time ![]() . If no further
input is applied for t >
. If no further
input is applied for t > ![]() , the voltage trajectory will have a pulse-like
excursion before it eventually returns to the resting potential. For
t >
, the voltage trajectory will have a pulse-like
excursion before it eventually returns to the resting potential. For
t > ![]() , we may therefore set
u(t) =
, we may therefore set
u(t) = ![]() (t -
(t - ![]() ) + urest where
) + urest where
![]() is the standard shape of the pulse and
urest is the resting
potential. We have
is the standard shape of the pulse and
urest is the resting
potential. We have
![]() (t -
(t - ![]() )
)![]() 0 for
t -
0 for
t - ![]()
![]()
![]() , because,
without further input, the voltage will eventually approach the resting value.
, because,
without further input, the voltage will eventually approach the resting value.
Let us now consider an additional small input current pulse I which is
applied at t > ![]() . Due to the input, the membrane potential will be
slightly perturbed from its trajectory. If the input current is sufficiently
small, the perturbation can be described by a linear impulse response function
. Due to the input, the membrane potential will be
slightly perturbed from its trajectory. If the input current is sufficiently
small, the perturbation can be described by a linear impulse response function
![]() . The response to an input pulse, and therewith the response
kernel
. The response to an input pulse, and therewith the response
kernel ![]() , can depend on the
arrival time of the input relative to the last spike at
, can depend on the
arrival time of the input relative to the last spike at ![]() .
For an input with arbitrary
time course I(t) we therefore set
.
For an input with arbitrary
time course I(t) we therefore set
To construct an approximative mapping between the SRM (4.63) and the
Hodgkin-Huxley equations, we have to determine the following three terms:
(i) the kernel ![]() which describes the response to spike emission, (ii) the
kernel
which describes the response to spike emission, (ii) the
kernel ![]() which describes the response to incoming current, and (iii)
the value of the threshold
which describes the response to incoming current, and (iii)
the value of the threshold ![]() .
.
In the absence of input the membrane potential u is at its resting value
urest. If we apply a strong current pulse, an action potential will
be triggered. The time course of the action potential determines the kernel
![]() .
.
To find the kernel ![]() we use the following procedure. We take a square
current pulse of the form
we use the following procedure. We take a square
current pulse of the form
![\begin{minipage}{0.43\textwidth}
{\bf A}
\par\includegraphics[height=40mm,width...
...\includegraphics[height=40mm,width=\textwidth]{HH-epskappa6.eps}
\end{minipage}](img521.gif) |
The kernel ![]() characterizes the linear response of the neuron to a
weak input current pulse. To measure
characterizes the linear response of the neuron to a
weak input current pulse. To measure ![]() we use a first strong pulse
to initiate a spike at a time
we use a first strong pulse
to initiate a spike at a time
![]() < 0 and then apply a second weak pulse
at t = 0. The second pulse is a short stimulus
as in Eq. (4.64),
but with a small amplitude so that nonlinear effects in the response can be
neglected. The result is a membrane potential with time course u(t).
Without the second pulse the time course of the potential would be
u0(t) =
< 0 and then apply a second weak pulse
at t = 0. The second pulse is a short stimulus
as in Eq. (4.64),
but with a small amplitude so that nonlinear effects in the response can be
neglected. The result is a membrane potential with time course u(t).
Without the second pulse the time course of the potential would be
u0(t) = ![]() (t -
(t - ![]() ) + urest for t >
) + urest for t > ![]() . The net effect of the second
pulse is
u(t) - u0(t), hence
. The net effect of the second
pulse is
u(t) - u0(t), hence
The result is shown in Fig. 4.12. Since the input current pulse
delivers its charge during a very short amount of time,
the ![]() -kernel jumps instantaneously at time t = 0 to a value
of 1mV. Afterwards it decays, with a slight oscillation, back to zero.
The oscillatory behavior is characteristic for
type II neuron models (Izhikevich, 2001).
The
decay of the oscillation
is faster if there has been a spike in the recent past. This is easy to
understand intuitively. During and immediately after an action potential many
ion channels are open. The resistance of the cell membrane is therefore
reduced and the effective membrane time constant is shorter;
cf. Fig. 2.7B.
-kernel jumps instantaneously at time t = 0 to a value
of 1mV. Afterwards it decays, with a slight oscillation, back to zero.
The oscillatory behavior is characteristic for
type II neuron models (Izhikevich, 2001).
The
decay of the oscillation
is faster if there has been a spike in the recent past. This is easy to
understand intuitively. During and immediately after an action potential many
ion channels are open. The resistance of the cell membrane is therefore
reduced and the effective membrane time constant is shorter;
cf. Fig. 2.7B.
The third term to be determined is the threshold ![]() which we will
take as fixed. Even though Fig. 2.4B suggests that the
Hodgkin-Huxley equations exhibit a certain form of threshold behavior, the
threshold is not well-defined (Koch et al., 1995; Rinzel and Ermentrout, 1989)
and it is fairly difficult to estimate a voltage threshold directly from a
single series of simulations. We therefore take the threshold as a free
parameter which will be adjusted by a procedure discussed below.
which we will
take as fixed. Even though Fig. 2.4B suggests that the
Hodgkin-Huxley equations exhibit a certain form of threshold behavior, the
threshold is not well-defined (Koch et al., 1995; Rinzel and Ermentrout, 1989)
and it is fairly difficult to estimate a voltage threshold directly from a
single series of simulations. We therefore take the threshold as a free
parameter which will be adjusted by a procedure discussed below.
In order to test the fidelity of the Spike Response Model we use the same input scenarios as in Chapter 2.2 for the Hodgkin-Huxley model. In particular, we consider constant input current, step current, and flucutating input current. We start with the time-dependent fluctuating input, since this is probably the most realistic scenario. We will see that the Spike Response Model with the kernels that have been derived above can approximate the spike train of the Hodgkin-Huxley model to a high degree of accuracy.
To test the quality of the SRM approximation we compare the spike trains
generated by the Spike Response Model (4.63) with that of the full
Hodgkin-Huxley model (2.4)-(2.6). We study the case of a
time-dependent input current I(t) generated by the procedure discussed in
section 2.2.2; cf. Fig. 2.7. The same current
is applied to both the Hodgkin-Huxley and the Spike Response model. The
threshold ![]() of the Spike Response Model has been adjusted so that
the total number of spikes was about the same as in the Hodgkin-Huxley model;
see Kistler et al. (1997) for details. In Fig. 4.13 the voltage trace of
the Hodgkin-Huxley model is compared to that of the Spike Response Model with
the kernels
of the Spike Response Model has been adjusted so that
the total number of spikes was about the same as in the Hodgkin-Huxley model;
see Kistler et al. (1997) for details. In Fig. 4.13 the voltage trace of
the Hodgkin-Huxley model is compared to that of the Spike Response Model with
the kernels ![]() and
and ![]() derived above. We see that the
approximation is excellent both in the absence of spikes and during spiking.
As an aside we note that it is indeed important to include the dependence of
the kernel
derived above. We see that the
approximation is excellent both in the absence of spikes and during spiking.
As an aside we note that it is indeed important to include the dependence of
the kernel ![]() upon the last output spike time
upon the last output spike time ![]() . If we
neglected that dependence and used
. If we
neglected that dependence and used
![]() (
(![]() , s) instead of
, s) instead of
![]() (t -
(t - ![]() , s), then the approximation during and immediately after a
spike would be significantly worse; see the dotted line in the lower right
graph of Fig. 4.13.
, s), then the approximation during and immediately after a
spike would be significantly worse; see the dotted line in the lower right
graph of Fig. 4.13.
![\centerline{\includegraphics[width=95mm]{HH-approx.ps}}](img525.gif) |
To check whether both models generated spikes at the same time we introduce the coincidence rate
We find that the Spike Response Model (4.63) reproduces the firing times
and the voltage time course of the Hodgkin-Huxley model to a high degree of
accuracy; cf. Fig. 4.13. More precisely, the Spike Response Model
achieves with a fluctuating input current a coincidence rate ![]() of about
0.85 (Kistler et al., 1997). On the other hand, a leaky integrate-and-fire model
with optimized time constant and fixed threshold yields coincidence rates in
the range of only 0.45. The difference in the performance of Spike Response
and integrate-and-fire model is not too surprising because the Spike Response
Model accounts for the hyperpolarizing spike after-potential; cf.
Fig 4.12A. In fact, an integrate-and-fire model with spike
after-potential (or equivalently a dynamic threshold) achieves coincidence
rates in the range of
of about
0.85 (Kistler et al., 1997). On the other hand, a leaky integrate-and-fire model
with optimized time constant and fixed threshold yields coincidence rates in
the range of only 0.45. The difference in the performance of Spike Response
and integrate-and-fire model is not too surprising because the Spike Response
Model accounts for the hyperpolarizing spike after-potential; cf.
Fig 4.12A. In fact, an integrate-and-fire model with spike
after-potential (or equivalently a dynamic threshold) achieves coincidence
rates in the range of
![]()
![]() 0.7 (Kistler et al., 1997). Furthermore, the
0.7 (Kistler et al., 1997). Furthermore, the
![]() -kernel of the Spike Response Model describes the reduced
responsiveness of the Hodgkin-Huxley model immediately after a spike; cf.
Fig 4.12B. The model SRM0 (with a kernel
-kernel of the Spike Response Model describes the reduced
responsiveness of the Hodgkin-Huxley model immediately after a spike; cf.
Fig 4.12B. The model SRM0 (with a kernel ![]() that
does not depend on t -
that
does not depend on t - ![]() ) yields a coincidence rate
) yields a coincidence rate ![]() that is
significantly lower than that of the full Spike Response Model.
that is
significantly lower than that of the full Spike Response Model.
We study the response of the Spike Response Model to constant stimulation
using the kernels derived by the procedure described above. The result is
shown in Fig. 4.14. As mentioned above, we take the threshold
![]() as a free parameter. If
as a free parameter. If ![]() is optimized for stationary
input, the frequency plots of the Hodgkin-Huxley model and the Spike Response
Model are rather similar. On the other hand, if we took the value of the
threshold that was found for time-dependent input, the current threshold for
the Spike Response Model would be quite different as shown by the dashed line
in Fig. 4.14.
is optimized for stationary
input, the frequency plots of the Hodgkin-Huxley model and the Spike Response
Model are rather similar. On the other hand, if we took the value of the
threshold that was found for time-dependent input, the current threshold for
the Spike Response Model would be quite different as shown by the dashed line
in Fig. 4.14.
![\centerline{\includegraphics[width=55mm]{HH-gain-all.eps} }](img533.gif) |
As a third input paradigm, we test the Spike Response Model with step current
input. For ![]() we take the value found for the scenario with
time-dependent input. The result is shown in Fig. 4.15. The
Spike Response Model shows the same three regimes as the Hodgkin-Huxley model.
In particular, the effect of inhibitory rebound is present in the Spike
Response Model. The location of the phase boundaries depends on the choice of
we take the value found for the scenario with
time-dependent input. The result is shown in Fig. 4.15. The
Spike Response Model shows the same three regimes as the Hodgkin-Huxley model.
In particular, the effect of inhibitory rebound is present in the Spike
Response Model. The location of the phase boundaries depends on the choice of
![]() .
.
![\centerline{\includegraphics[width=120mm]{Fig10-HH-SRM-phase2.eps}}](img534.gif) |
In the Hodgkin-Huxley model (2.4), input is formulated as an explicit
driving current I(t). In networks of neurons, input typically consists of
the spikes of presynaptic neurons. Let us, for the sake of
simplicity, assume that a spike of a presynaptic neuron j which was emitted
at time tj(f) generates in the postsynaptic neuron i a current
I(t) = wij ![]() (t - tj(f)). Here,
(t - tj(f)). Here, ![]() describes the time course of the
postsynaptic current and wij scales the amplitude of the current. The
voltage of the postsynaptic neuron i changes, according to (4.63) by
an amount
describes the time course of the
postsynaptic current and wij scales the amplitude of the current. The
voltage of the postsynaptic neuron i changes, according to (4.63) by
an amount
![]() ui(t) = wij
ui(t) = wij ![]()
![]() (t -
(t - ![]() , s)
, s) ![]() (t - tj(f) - s) ds, where
(t - tj(f) - s) ds, where ![]() is the last output spike of
neuron i. The voltage response
is the last output spike of
neuron i. The voltage response
![]() ui to an input current of unit
amplitude (wij = 1) defines the postsynaptic potential
ui to an input current of unit
amplitude (wij = 1) defines the postsynaptic potential ![]() , hence
, hence
We have seen in the previous section that the Spike Response Model can provide a good quantitative approximation of the Hodgkin-Huxley model. Though the Hodgkin-Huxley equation captures the essence of spike generation it is ``only'' a model of the giant axon of the squid which has electrical properties that are quite different from those of cortical neurons we are mostly interested in. The natural question is thus whether the Spike Response Model can also be used as a quantitative model of cortical neurons. In the following we discuss a conductance-based neuron model for a cortical interneuron and show how such a model can be reduced to a (nonlinear) integrate-and-fire model or to a Spike Response Model.
The starting point is a conductance-based model that has originally been proposed as a model of fast-spiking neo-cortical interneurons (Erisir et al., 1999). We have chosen this specific model for two reasons: First, just as most other cortical neuron models, this model has - after a minor modification - a continuous gain function (Lewis and Gerstner, 2001) and can hence be classified as a type I model (Ermentrout, 1996). This is in contrast to the Hodgkin-Huxley model which exhibits a discontinuity in the gain function and is hence type II. Second, this is a model for interneurons that show little adaptation, so that we avoid most of the complications caused by slow ionic processes that cannot be captured by the class of spiking neuron models reviewed above. Furthermore, the model is comparatively simple, so that we can hope to illustrate the steps necessary for a reduction to formal spiking neuron models in a transparent manner.
The model neuron consists of a single compartment with a non-specific leak current and three types of ion current, i.e., a Hodgkin-Huxley type sodium current INa = gNa m3 h (u - ENa), a slow potassium current Islow = gKslow n4slow (u - EK), and a fast potassium current Ifast = gKfast n2fast (u - EK). The response properties of the cortical neuron model to pulse input and constant current have already been discussed in Chapter 2.3; cf. Fig. 2.11. We now want to reduce the model to a nonlinear integrate-and fire model or, alternatively, to a Spike Response Model. We start with the reduction to an integrate-and-fire model.
In order to reduce the dynamics of the full cortical neuron model to that of an integrate-and-fire model we proceed in two steps. As a first step, we keep all variables, but introduce a threshold for spike initiation. We call this the multi-current integrate-and-fire model. In the second step, we separate gating variables into fast and slow ones. `Fast' variables are replaced by their steady state values, while `slow' variables are replaced by constants. The result is the desired nonlinear integrate-and-fire model with a single dynamical variable.
In step (i), we make use of the observation that the shape of an action
potential of the cortical neuron model is always roughly the same,
independently of the way the spike is initiated; cf.
Fig. 4.16. Instead of calculating the shape of an action
potential again and again, we can therefore simply stop the costly numerical
integration of the nonlinear differential equations as soon as a spike is
triggered and restart the integration after the down-stroke of the spike about
1.5-2ms later. We call such a scheme a multi-current integrate-and-fire
model. The interval between the spike trigger time ![]() and the restart of
the integration corresponds to an absolute refractory period
and the restart of
the integration corresponds to an absolute refractory period
![]() .
.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=50mm]{A0-e...
...h}
{\bf B}
\par\includegraphics[height=50mm]{A0-h-compared.eps}
\end{minipage}](img537.gif) |
In order to transform the cortical neuron model into a multi-current
integrate-and-fire model we have to define a voltage threshold ![]() , a
refractory time
, a
refractory time
![]() , and the reset values from which the integration
is restarted. We fix the threshold at
, and the reset values from which the integration
is restarted. We fix the threshold at
![]() = - 40 mV; the exact value
is not critical and we could take values of -20mV or -45mV without
changing the results. For
= - 40 mV; the exact value
is not critical and we could take values of -20mV or -45mV without
changing the results. For
![]() = - 40 mV, a refractory time
= - 40 mV, a refractory time
![]() = 1.7ms and a reset voltage
ur = - 85 mV is suitable; cf. Fig. 4.16.
= 1.7ms and a reset voltage
ur = - 85 mV is suitable; cf. Fig. 4.16.
To restart the integration of the differential equation we also have to
specify initial conditions for the gating variables
m, h, nslow, and
nfast. This, however, involves a severe simplification, because their
time course is not as stereotyped as that of the membrane potential, but
depends on the choice of the input scenario; cf. Fig. 4.16B.
In the following we optimize the reset values for a scenario with a constant
input current
Iext = 5 ![]() A/cm2 that leads to repetitive firing
at about 40Hz. The reset values are
mr = 0.0;hr = 0.16;nslow, r = 0.874;nfast, r = 0.2; and
ur = - 85 mV. This
set of parameters yields a near-perfect fit of the time course of the membrane
potential during repetitive firing at 40Hz and approximates the gain
function of the full cortical neuron model to a high degree of accuracy; cf.
Fig. 4.17.
A/cm2 that leads to repetitive firing
at about 40Hz. The reset values are
mr = 0.0;hr = 0.16;nslow, r = 0.874;nfast, r = 0.2; and
ur = - 85 mV. This
set of parameters yields a near-perfect fit of the time course of the membrane
potential during repetitive firing at 40Hz and approximates the gain
function of the full cortical neuron model to a high degree of accuracy; cf.
Fig. 4.17.
![\centerline{\includegraphics[width=6cm]{A1gain.eps}}](img539.gif) |
So far the model contains still all five variables u, m, h, nslow, nfast. To get rid of all the gating variables we distinguish between variables x that are either fast as compared to u - in which case we replace it by its steady-state value x0(u) - or slow as compared to u - in which case we replace x by a constant. Here, m is the only fast variable and we replace m(t) by its steady-state value m0[u(t)]. The treatment of the other gating variables deserves some extra discussion.
A thorough inspection of the time
course of
nfast(t) shows that
nfast is most of the time
close to its resting value, except for a 2ms interval during and immediately
after the down-stroke of an action potential. If we take a refractory time of
![]() = 4 ms, most of the excursion trajectory of
nfast falls
within the refractory period. Between spikes we can therefore replace
nfast by its equilibrium value at rest
nfast, rest = n0, fast(urest).
= 4 ms, most of the excursion trajectory of
nfast falls
within the refractory period. Between spikes we can therefore replace
nfast by its equilibrium value at rest
nfast, rest = n0, fast(urest).
The gating variables h and nslow vary slowly, so that the variables may be replaced by averaged values hav and nslow, av. The average, however, depends on the input scenario. We stick to a regime with repetitive firing at 40Hz where hav = 0.7 and nslow, av = 0.8.
With m = m0(u) and constant values for h, nslow, nfast, the dynamics of the full cortical neuron model reduces to
| C |
= gNa [m0(u)]3 hav (u - ENa) + gKslow nslow, av2 (u - EK) | |
| + gKfast nfast, rest4 (u - EK) + gl (u - El) + Iext(t). | (4.70) |
The passive membrane time constant of the model is inversely proportional to
the slope of F at rest:
![]() = | dF/du|-1u=urest. In principle the function F could be
further approximated by a linear function with slope -1/
= | dF/du|-1u=urest. In principle the function F could be
further approximated by a linear function with slope -1/![]() and then
combined with a threshold at, e.g.,
and then
combined with a threshold at, e.g.,
![]() = - 45 mV. This would yield a
linear integrate-and-fire model. Alternatively, F can be
approximated by a quadratic function which leads us to a quadratic
integrate-and-fire neuron; cf. Fig. 4.18.
= - 45 mV. This would yield a
linear integrate-and-fire model. Alternatively, F can be
approximated by a quadratic function which leads us to a quadratic
integrate-and-fire neuron; cf. Fig. 4.18.
![\centerline{\includegraphics[width=6cm]{A3-current-quad.eps}}](img540.gif) |
To test the fidelity of the reduction to a nonlinear integrate-and-fire model, we compare its behavior to that of the full cortical neuron model for various input scenarios. It turns out that the behavior of the model is good as long as the mean firing rate is in the range of 40Hz, which is not too surprising given the optimization of the parameters for this firing rate. Outside the range of 40±10Hz there are substantial discrepancies between the reduced and the full model.
Let us focus on constant input first. With our set of parameters
we get a fair approximation of the gain function, except that the
threshold for repetitive firing is not reproduced correctly; cf.
Fig. 4.19A. We note that the firing rate at a stimulation
of 5![]() A/cm2 is reproduced correctly which is no surprise since our
choice of parameters has been based on this input amplitude.
A/cm2 is reproduced correctly which is no surprise since our
choice of parameters has been based on this input amplitude.
For a critical test of the nonlinear integrate-and-fire model, we use a
fluctating input current with zero mean. The amplitude of the fluctuations
determines the mean firing rate. The nonlinear integrate-and-fire model,
however, does not reproduce the firing rate as a function of the fluctuation
amplitude of the full model, except at
![]()
![]() 40Hz; cf. Figure 4.19B.
40Hz; cf. Figure 4.19B.
For a more detailed comparison of the nonlinear integrate-and-fire with the
full model, we stimulate both models with the same fluctuating current. From
Fig. 4.20A, we see that the voltage time course of the two
models is most of the time indistinguishable. Occasionally, the nonlinear
integrate-and-fire model misses a spike, or adds an extra spike. For this
specific input scenario (where the mean firing rate is about 40Hz), a
coincidence rate of about ![]() = 85 is achieved (based on a precision of
= 85 is achieved (based on a precision of
![]() = ±2ms). Outside the regime of
= ±2ms). Outside the regime of
![]()
![]() 40Hz, the
coincidence rate
40Hz, the
coincidence rate ![]() breaks down drastically; cf.
Fig. 4.20B.
breaks down drastically; cf.
Fig. 4.20B.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
...{\bf B}
\par\includegraphics[width=\textwidth]{A3-ratesrefr.eps}
\end{minipage}](img541.gif) |
As a second approximation scheme, we consider the reduction of the
conductance-based neuron model to a Spike Response Model. We thus have to
determine the kernels
![]() (t -
(t - ![]() ),
),
![]() (t -
(t - ![]() , s), and adjust the
(time-dependent) threshold
, s), and adjust the
(time-dependent) threshold
![]() (t -
(t - ![]() ). We proceed in three steps.
As a first step we reduce the model to an integrate-and-fire model with
spike-time dependent time constant. As a second step, we integrate the model
so as to derive the kernels
). We proceed in three steps.
As a first step we reduce the model to an integrate-and-fire model with
spike-time dependent time constant. As a second step, we integrate the model
so as to derive the kernels ![]() and
and ![]() . As a final step, we
choose an appropriate spike-time dependent threshold.
. As a final step, we
choose an appropriate spike-time dependent threshold.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
...B}
\par\includegraphics[width=0.9\textwidth]{A1-A3-A4-gamma.eps}
\end{minipage}](img542.gif) |
In step (i), we stimulate the full model by a short
super-threshold current pulse in order to determine the time course of the
action potential and its hyper-polarizing spike after-potential. Let
us define ![]() as the time when the membrane potential crosses an
(arbitrarily fixed) threshold
as the time when the membrane potential crosses an
(arbitrarily fixed) threshold ![]() , e.g.,
, e.g.,
![]() = - 50 mV. The
time course of the membrane potential for t >
= - 50 mV. The
time course of the membrane potential for t > ![]() , i.e., during and after
the action potential, defines the kernel
, i.e., during and after
the action potential, defines the kernel
![]() (t -
(t - ![]() ). If we were
interested in a purely phenomenological model, we could simply record the
numerical time course u(t) and define
). If we were
interested in a purely phenomenological model, we could simply record the
numerical time course u(t) and define
![]() (t -
(t - ![]() ) = u(t) - urest
for t >
) = u(t) - urest
for t > ![]() ; cf. Section 4.3.1. It is, however,
instructive to take a semi-analytical approach and study the four gating
variables
m, h, nslow and
nfast. About 2ms after initiation
of the spike, all four variables have passed their maximum or minimal values
and are on their way back to equilibrium.
We set
; cf. Section 4.3.1. It is, however,
instructive to take a semi-analytical approach and study the four gating
variables
m, h, nslow and
nfast. About 2ms after initiation
of the spike, all four variables have passed their maximum or minimal values
and are on their way back to equilibrium.
We set
![]() = 2ms. For
t
= 2ms. For
t![]()
![]() +
+ ![]() , we fit the approach
to equilibrium by an exponential
, we fit the approach
to equilibrium by an exponential
Given the time course of the gating variables, we know the conductance of each ion channel as a function of time. For example, the potassium current IKfast is
In step (ii) we integrate Eq. (4.75) with the initial condition
| u( |
(4.76) |
u(t) = 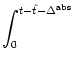 |
(4.77) |
![\centerline{\includegraphics[width=6cm]{A2-tau.eps}}](img554.gif) |
Finally, in step (iii) we introduce a dynamical threshold
We now test the Spike Response Model with the above set of parameters on a
scenario with fluctuating input current. The mean firing rate of the full
cortical neuron model and the Spike Response Model as a function of the
fluctuation amplitude are similar; cf. Fig. 4.22B.
Moreover, there is a high percentage of firing times of the Spike Response
Model that coincide with those of the full model with a precision of
![]() = ±2ms [coincidence rate
= ±2ms [coincidence rate
![]() = 0.75; cf. Eq. (4.67)]. A sample
spike train is shown in Fig. 4.23A. Figure
= 0.75; cf. Eq. (4.67)]. A sample
spike train is shown in Fig. 4.23A. Figure ![]() B exhibits a plot of the coincidence measure
B exhibits a plot of the coincidence measure ![]() defined in equation
(4.67) as a function of the fluctuation amplitude. In
contrast to the nonlinear integrate-and-fire neuron, the coincidence rate is
fairly constant over a broad range of stimulus amplitudes. At low rates,
however, the coincidence rate drops off rapidly.
defined in equation
(4.67) as a function of the fluctuation amplitude. In
contrast to the nonlinear integrate-and-fire neuron, the coincidence rate is
fairly constant over a broad range of stimulus amplitudes. At low rates,
however, the coincidence rate drops off rapidly.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
...{\bf B}
\par\includegraphics[width=\textwidth]{A4-randrates.eps}
\end{minipage}](img559.gif) |
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
...par\includegraphics[width=0.9\textwidth]{A1-A3-A4-gamma-SRM.eps}
\end{minipage}](img560.gif) |
Not surprisingly, each approximation scheme is only valid in a limited regime.
The natural question is thus whether this is the biologically relevant regime.
Since a fluctuating input is probably the most realistic scenario, we have
focused our discussion on this form of stimulation. We have seen that in
case of a fluctuating input current, integrate-and-fire and Spike Response
Model reproduce - to a certain extend - not only the mean
firing rate, but also the firing times of the corresponding detailed neuron
model. In this discipline, the multi-current integrate-and-fire
model clearly yields the best performance. While it is easy to implement and
rapid to simulate, it is difficult to analyze mathematically. Strictly
speaking, it does not fall in the class of spiking neuron models reviewed in
this chapter. It is interesting to see, however, that even the
multi-current integrate-and-fire model which is based on a seemingly innocent
approximation exhibits, for time-dependent input, a coincidence rate ![]() significantly below one. On the fluctuating-input task, we find that the
single-variable (nonlinear) integrate-and-fire model exhibits a pronounced
peak of
significantly below one. On the fluctuating-input task, we find that the
single-variable (nonlinear) integrate-and-fire model exhibits a pronounced
peak of ![]() at the optimal input, but does badly outside this regime. A
Spike Response Model without adapting threshold yields coincidence
rates that are not significantly different from the results for the nonlinear
integrate-and-fire model. This indicates that the time-dependent threshold
that has been included the definition of the Spike Response Model is an
important component to achieve generalization over a broad range of different
inputs. This also suggests that the nonlinear dependence of F(u) upon the
membrane potential is not of eminent importance for the random-input task.
at the optimal input, but does badly outside this regime. A
Spike Response Model without adapting threshold yields coincidence
rates that are not significantly different from the results for the nonlinear
integrate-and-fire model. This indicates that the time-dependent threshold
that has been included the definition of the Spike Response Model is an
important component to achieve generalization over a broad range of different
inputs. This also suggests that the nonlinear dependence of F(u) upon the
membrane potential is not of eminent importance for the random-input task.
On the other hand, in the immediate neighborhood of the firing threshold, the nonlinear integrate-and-fire model performs better than the Spike Response Model. In fact, the Spike Response Model systematically fails to reproduce delayed action potentials triggered by input that is just slightly super-threshold. As we have seen, the nonlinear integrate-and-fire model is related to a canonical type I model and, therefore, exhibits the `correct' behavior in the neighborhood of the firing threshold.
In summary, it is always possible to design an input scenario where formal spiking neuron models fail. For example, none of the models discussed in this chapter is capable of reproducing effects of a slow adaptation to changes in the input.
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
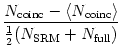

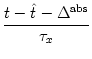
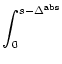 exp
exp![$\displaystyle \begin{array}{*{2}{c@{\qquad}}c} \vartheta^{\rm refr} &{\rm for} ...
... \tau_\vartheta} \right) \right] &{\rm for} &s \ge \Delta^{\rm abs} \end{array}$](img556.gif)