In many areas of the brain neurons are organized in populations of units with
similar properties. Prominent examples are columns in the somatosensory and
visual cortex (Hubel and Wiesel, 1962; Mountcastle, 1957) and pools of motor neurons
(Kandel and Schwartz, 1991). Given the large number of neurons within such a column or
pool it is sensible to describe the mean activity of the neuronal population
rather than the spiking of individual neurons. The idea of a population
activity has already been introduced in Chapter 1.4. In a
population of N neurons, we calculate the proportion of active neurons by
counting the number of spikes
nact(t;t + ![]() t) in a small time
interval
t) in a small time
interval ![]() t and dividing by N. Further division by
t and dividing by N. Further division by ![]() t
yields the population activity
t
yields the population activity
Theories of population activity have a long tradition (Nykamp and Tranchina, 2000; Treves, 1993; Wilson and Cowan, 1972; Amari, 1974; Wilson and Cowan, 1973; Gerstner and van Hemmen, 1992; Brunel, 2000; Abbott and van Vreeswijk, 1993; Gerstner, 1995; Fusi and Mattia, 1999; Brunel and Hakim, 1999; Gerstner, 2000b; Amit and Brunel, 1997a,b; Eggert and van Hemmen, 2001; Knight, 1972a; Omurtag et al., 2000). In this chapter we study the properties of a large and homogeneous population of spiking neurons. Why do we restrict ourselves to large populations? If we repeatedly conduct the same experiment on a population of, say, one hundred potentially noisy neurons, the observed activity A(t) defined in Eq. (6.1) will vary from one trial to the next. Therefore we cannot expect a population theory to predict the activity measurements in a single trial. Rather all population activity equations that we discuss in this chapter predict the expected activity. For a large and homogeneous network, the observable activity is very close to the expected activity. For the sake of notational simplicity, we do not distinguish the observed activity from its expectation value and denote in the following the expected activity by A(t).
After clarifying the notion of a homogeneous network in Section 6.1, we derive in Section 6.2 population density equations, i.e., partial differential equations that describe the probability that an arbitrary neuron in the population has a specific internal state. In some special cases, these density equations can be integrated and presented in the form of an integral equation. In Section 6.3 a general integral equation for the temporal evolution of the activity A(t) that is exact in the limit of a large number of neurons is derived. In particular, we discuss its relation to the Wilson-Cowan equation, one of the standard models of population activity. In Section 6.4 we solve the population equation for the fixed points of the population activity and show that the neuronal gain function plays an important role. Finally, in Section 6.5 the approach is extended to multiple populations and its relation to neuronal field equations is discussed.
Most of the discussion in part II of the present book will be based upon the population equations introduced in this chapter. The population activity equations will allow us to study signal transmission and coding (cf. Chapter 7), oscillations and synchronization (cf. Chapter 8), and the formation of activity patterns in populations with a spatial structure (cf. Chapter 9). The aim of the present chapter is two-fold. Firstly, we want to provide the reader with the mathematical formalism necessary for a systematic study of spatial and temporal phenomena in large populations of neurons. Secondly, we want to show that various formulations of population dynamics that may appear quite different at a first glance, are in fact closely related. Paragraphs that are more mathematically oriented are marked by an asterix and can be omitted at a first reading.
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
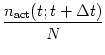 =
=