We define asynchronous firing of a neuronal population as a macroscopic firing state with constant activity A(t) = A0. In this section we use the population activity equations (6.73) and (6.75) to study the existence of asynchronous firing states in a homogeneous population of spiking neurons. We will see that the neuronal gain function plays an important role. More specifically, we will show that the knowledge of the single-neuron gain function g(I0) and the coupling parameter J0 is sufficient to determine the activity A0 during asynchronous firing.
![\hbox{{\bf A} \hspace{62mm} {\bf B}}
\hbox{\hspace{10mm}
\includegraphics[width=...
...% includegraphics[width=50mm]\{Figs-ch-pop/sp-Aoft-large-4.eps\}\}
\vspace{0mm}](img1108.gif) |
In this section we will show that
during asynchronous firing the population activity A0
is equal to the mean firing rate of a single neuron
in the population. To do so, we
search for a stationary solution A(t) = A0 of the
population equation (6.73). Given constant activity
A0
and constant external input
Iext0,
the total input I0 to each neuron is constant.
In this case, the state of each neuron
depends only on t - ![]() , i.e.,
the time since its last output spike.
We are thus in the situation
of stationary renewal theory.
, i.e.,
the time since its last output spike.
We are thus in the situation
of stationary renewal theory.
In the stationary state, the survivor function
and the interval distribution
can not depend explicitly upon
the absolute time, but only on
the time difference
s = t - ![]() .
Hence we set
.
Hence we set
| SI( |
S0(s) | (6.92) | |
| PI( |
P0(s) | (6.93) |
| 1 = A0 |
(6.95) |
| (6.96) |
![\hbox{{\bf A} \hspace{65mm} {\bf B}}
\hbox{\hspace{10mm}
\includegraphics[width=...
...\includegraphics[width=53mm]{Figs-ch-pop/spikeraster-nonoise.eps}}
\vspace{0mm}](img1110.gif) |
How can we compare the population activity A0 calculated in
Eq. (6.97) with simulation results? In a simulation of a
population containing a finite number N of spiking neurons, the observed
activity fluctuates. Formally, the (observable) activity A(t) has been
defined in Eq. (6.1) as a sum over ![]() functions. The
activity A0 predicted by the theory is the expectation value of
the observed activity. Mathematically speaking, the observed activity A
converges for
N
functions. The
activity A0 predicted by the theory is the expectation value of
the observed activity. Mathematically speaking, the observed activity A
converges for
N![]()
![]() in the weak topology to its expectation value.
More practically this implies that we should convolve the observed activity
with a continuous test function
in the weak topology to its expectation value.
More practically this implies that we should convolve the observed activity
with a continuous test function ![]() (s) before comparing with A0. We
take a function
(s) before comparing with A0. We
take a function ![]() with the normalization
with the normalization
![]()
![]() (s) ds = 1. For the sake of simplicity we assume furthermore that
(s) ds = 1. For the sake of simplicity we assume furthermore that
![]() has finite support so that
has finite support so that
![]() (s) = 0 for s < 0 or
s > smax. We define
(s) = 0 for s < 0 or
s > smax. We define
| (6.98) |
For the purpose of illustration, we have plotted
in Fig. 6.10A the spikes of
eight neurons of the network simulation shown in
Fig. 6.9. The mean
interspike-interval for a single neuron is
![]() T
T![]() = 20ms
which corresponds to a population activity
of A0 = 50Hz.
= 20ms
which corresponds to a population activity
of A0 = 50Hz.
The gain function of a neuron is the firing rate
![]() T
T![]() as a
function of its input current I. In the previous subsection, we have seen
that the firing rate is equivalent to the population activity A0 in the
state of asynchronous firing. We thus have
as a
function of its input current I. In the previous subsection, we have seen
that the firing rate is equivalent to the population activity A0 in the
state of asynchronous firing. We thus have
Figure 6.11 shows a graphical solution of Eq. (6.102) in terms
of the mean interval
![]() T
T![]() as a function of the input I0
(i.e., the gain function) and the total input I0 as a function of the
activity A0. The intersections of the two functions yield fixed points of
the activity A0.
as a function of the input I0
(i.e., the gain function) and the total input I0 as a function of the
activity A0. The intersections of the two functions yield fixed points of
the activity A0.
![\hbox{\hspace{20mm}
\includegraphics[width=50mm]{Figs-ch-pop/fig15b.eps}}](img1117.gif) |
As an aside we note that the graphical construction is identical to that of the Curie-Weiss theory of ferromagnetism which can be found in any physics textbook. More generally, the structure of the equations corresponds to the mean-field solution of a system with feedback. As shown in Fig. 6.11, several solutions may coexist. We cannot conclude from the figure, whether one or several solutions are stable. In fact, it is possible that all solutions are unstable. In the latter case, the network leaves the state of asynchronous firing and evolves towards an oscillatory or quiescent state. The stability analysis of the asynchronous state is deferred to Chapter 8.
Consider a population of (noisy) SRM0 neurons with escape rate f,
e.g.
f (u - ![]() ) = exp[
) = exp[![]() (u -
(u - ![]() )];
cf. Chapter 5.3. The stationary activity A0 in the presence
of a constant input potential
h0 = R I0 is given by
)];
cf. Chapter 5.3. The stationary activity A0 in the presence
of a constant input potential
h0 = R I0 is given by
In the limit of diffusive noise the stationary activity is
| I0 = Iext0 + J0 A0 ; | (6.105) |
![\begin{minipage}{0.4\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]{...
...ar\includegraphics[width=\textwidth]{Figs-ch-pop/gain-IF-s0.eps}
\end{minipage}](img1127.gif) |
In the preceding subsections we have studied the stationary state of a population of neurons for a given noise level. The noise was modeled either as diffusive noise mimicking stochastic spike arrival or as escape noise mimicking a noisy threshold. In both cases noise was added explicitly to the model. In this section we discuss how a network of deterministic neurons with fixed random connectivity can generate its own noise. In particular, we will focus on spontaneous activity and argue that there exist stationary states of asynchronous firing at low firing rates which have broad distributions of interspike intervals even though individual neurons are deterministic. This point has been emphasized by van Vreeswijk and Sompolinsky (1996,1998) who used a network of binary neurons to demonstrate broad interval distribution in deterministic networks. Amit and Brunel (1997a,b) where the first to analyze a network of integrate-and-fire neurons with fixed random connectivity. While they allowed for an additional fluctuating input current, the major part of the fluctuations were in fact generated by the network itself. The theory of randomly connected integrate-and-fire neurons has been further developped by Brunel and Hakim (1999). In a recent study, Brunel (2000) confirmed that asynchronous highly irregular firing can be a stable solution of the network dynamics in a completely deterministic network consisting of excitatory and inhibitory integrate-and-fire neurons. The analysis of randomly connected networks of integrate-and-fire neurons is closely related to earlier theories for random nets of formal analog or binary neurons (Nützel, 1991; Kree and Zippelius, 1991; Amari, 1977b,1972,1974; Crisanti and Sompolinsky, 1988; Cessac et al., 1994).
The network structure plays a central role in the arguments. While we assume that all neurons in the population are of the same type, the connectivity between the neurons in the population is not homogeneous. Rather it is random, but fixed. Each neuron in the population of N neurons receives input from C randomly selected neurons in the population. Sparse connectivity means that the ratio
As a consequence of the sparse random network connectivity two neurons i and
j share only a small number of common inputs. In the limit of C/N![]() 0
the probability that neurons i and j have a common presynaptic neuron
vanishes. Thus, if the presynaptic neurons fire stochastically, then
the input spike trains that arrive at neuron i and j are independent
(Kree and Zippelius, 1991; Derrida et al., 1987). In that case, the input of neuron i and j can be
described as stochastic spike arrival which, as we have seen, can be described
by a diffusive noise model.
0
the probability that neurons i and j have a common presynaptic neuron
vanishes. Thus, if the presynaptic neurons fire stochastically, then
the input spike trains that arrive at neuron i and j are independent
(Kree and Zippelius, 1991; Derrida et al., 1987). In that case, the input of neuron i and j can be
described as stochastic spike arrival which, as we have seen, can be described
by a diffusive noise model.
The above reasoning, however, is based on the assumption that the presynaptic neurons (that are part of the population) fire stochastically. To make the argument self-consistent, we have to show that the firing of the postsynaptic neuron is, to a good approximation, also stochastic. The self-consistent argument will be outlined in the following.
We have seen in Chapter 5 that integrate-and-fire neurons with diffusive noise generate spike trains with a broad distribution of interspike intervals when they are driven in the sub-threshold regime. We will use this observation to construct a self-consistent solution for the stationary states of asynchronous firing.
We consider two populations, an excitatory population with NE neurons and
an inhibitory population with NI neurons. We assume that excitatory and
inhibitory neurons have the same parameters ![]() ,
, ![]() , R, and
ur. In addition all neurons are driven a common external current
Iext. Each neuron in the population receives CE synapses from excitatory
neurons with weight wE > 0 and CI synapses from inhibitory neurons with
weight wI < 0. If an input spike arrives at the synapses of neuron i from
a presynaptic neuron j, its membrane potential changes by an amount
, R, and
ur. In addition all neurons are driven a common external current
Iext. Each neuron in the population receives CE synapses from excitatory
neurons with weight wE > 0 and CI synapses from inhibitory neurons with
weight wI < 0. If an input spike arrives at the synapses of neuron i from
a presynaptic neuron j, its membrane potential changes by an amount
![]() ui = wj where wj = wE if j is excitatory and wj = wI if j is
inhibitory. We set
ui = wj where wj = wE if j is excitatory and wj = wI if j is
inhibitory. We set
Since excitatory and inhibitory neurons receive
the same number of inputs in our model, we assume
that they fire with a common firing rate ![]() .
The total input potential generated by
the external current and by the lateral couplings
is
.
The total input potential generated by
the external current and by the lateral couplings
is
The arguments that have been developed above for low-connectivity networks can be generalized to fully connected networks with asymmetric random connectivity (Sompolinsky et al., 1988; van Vreeswijk and Sompolinsky, 1996; Ben Arous and Guionnet, 1995; Amari, 1972; Cessac et al., 1994).
In the preceding sections, we have often considered neurons driven by a mean
input potential h0 = 0.8 and a variance
![]() = 0.2. Let us find
connectivity parameters of our network so that
= 0.2. Let us find
connectivity parameters of our network so that
![]() = 0.2 is the result of
stochastic spike arrivals from presynaptic neurons within the network. As
always we set
R =
= 0.2 is the result of
stochastic spike arrivals from presynaptic neurons within the network. As
always we set
R = ![]() = 1 and
= 1 and
![]() = 10ms.
= 10ms.
Figure 6.13A shows that h0 = 0.8 and
![]() = 0.2
correspond to a firing rate of
A0 =
= 0.2
correspond to a firing rate of
A0 = ![]()
![]() 16 Hz.
We set wE = 0.025, i.e., 40 simultaneous spikes
are necessary to make a neuron fire.
Inhibition has the same strength
wI = - wE
so that g = 1.
We constrain our search to solutions with
CE = CI so that
16 Hz.
We set wE = 0.025, i.e., 40 simultaneous spikes
are necessary to make a neuron fire.
Inhibition has the same strength
wI = - wE
so that g = 1.
We constrain our search to solutions with
CE = CI so that
![]() = 1.
Thus, on the average, excitation and inhibition
balance each other. To get an average
input potential of h0 = 0.8 we need
therefore a constant driving current
Iext = 0.8.
= 1.
Thus, on the average, excitation and inhibition
balance each other. To get an average
input potential of h0 = 0.8 we need
therefore a constant driving current
Iext = 0.8.
To arrive at
![]() = 0.2 we solve Eq. (6.109) for
CE and find
CE = CI = 200. Thus for this choice of the parameters
the network generates enough noise to allow a stationary solution of
asynchronous firing at 16Hz.
= 0.2 we solve Eq. (6.109) for
CE and find
CE = CI = 200. Thus for this choice of the parameters
the network generates enough noise to allow a stationary solution of
asynchronous firing at 16Hz.
![\hbox{{\bf A} \hspace{58mm} {\bf B}}
\hbox{\hspace{10mm}
\includegraphics[width=...
...15mm}
\includegraphics[width=45mm]{Figs-ch-pop/gain-IF-s0.54.eps}}
\vspace{0mm}](img1134.gif) |
Note that, for the same parameter, the inactive state where all neurons are silent is also a solution. Using the methods discussed in this section we cannot say anything about the stability of these states. For the stability analysis see (Brunel, 2000) and Chapter 7.
About eighty percent of the neurons in the cerebral cortex
are excitatory and twenty percent inhibitory.
Let us suppose that we have NE = 8000
excitatory and NI = 2000 inhibitory neurons
in a cortical column. We assume random connectivity
and take
CE = 800,
CI = 200
so that
![]() = 1/4.
As before, excitatory synapses have
a weight
wE = 0.025, i.e,
an action potential can be triggered by the simultaneous arrival
of 40 presynaptic spikes.
If neurons are driven in the regime
close to threshold, inhibition
is rather strong and we take
wI = - 0.125 so that g = 5.
Even though we have less inhibitory than excitatory neurons,
the mean feedback is then dominated by
inhibition since
= 1/4.
As before, excitatory synapses have
a weight
wE = 0.025, i.e,
an action potential can be triggered by the simultaneous arrival
of 40 presynaptic spikes.
If neurons are driven in the regime
close to threshold, inhibition
is rather strong and we take
wI = - 0.125 so that g = 5.
Even though we have less inhibitory than excitatory neurons,
the mean feedback is then dominated by
inhibition since
![]() g > 1.
We search for a consistent solution
of Eqs. (6.108) - (6.110)
with a spontaneous activity of
g > 1.
We search for a consistent solution
of Eqs. (6.108) - (6.110)
with a spontaneous activity of ![]() = 8Hz.
= 8Hz.
Given the above parameters, the variance is
![]()
![]() 0.54;
cf. Eq. (6.109). The gain function of
integrate-and-fire neurons gives us for
0.54;
cf. Eq. (6.109). The gain function of
integrate-and-fire neurons gives us for ![]() = 8Hz a corresponding total
potential of
h0
= 8Hz a corresponding total
potential of
h0 ![]() 0.2; cf. Fig. 6.13B. To attain
h0 we have to apply an external stimulus
h0ext = R Iext
which is slightly larger than h0 since the net effect of the lateral
coupling is inhibitory. Let us introduce the effective coupling
Jeff =
0.2; cf. Fig. 6.13B. To attain
h0 we have to apply an external stimulus
h0ext = R Iext
which is slightly larger than h0 since the net effect of the lateral
coupling is inhibitory. Let us introduce the effective coupling
Jeff = ![]() CE wE (1 -
CE wE (1 - ![]() g). Using the above parameters we find
from Eq. (6.108)
h0ext = h0 - Jeff A0
g). Using the above parameters we find
from Eq. (6.108)
h0ext = h0 - Jeff A0 ![]() 0.6.
0.6.
The external input could, of course, be provided by (stochastic) spike arrival from other columns in the same or other areas of the brain. In this case Eq. (6.108) is to be replaced by
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
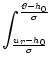 du exp
du exp