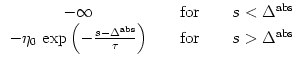There are various ways to introduce noise in formal spiking neuron models. In this section we focus on a `noisy threshold' (also called escape or hazard model). In section 5.5 we will discuss `noisy integration' (also called stochastic spike arrival or diffusion model). In both cases, we are interested in the effect of the noise on the distribution of interspike intervals.
In the escape model, we imagine that the neuron can fire even though the
formal threshold ![]() has not been reached or may stay quiescent
even though the formal threshold has been passed. To do this consistently, we
introduce an `escape rate' or `firing intensity' which depends on the
momentary state of the neuron.
has not been reached or may stay quiescent
even though the formal threshold has been passed. To do this consistently, we
introduce an `escape rate' or `firing intensity' which depends on the
momentary state of the neuron.
Given the input I and the firing time ![]() of the last spike, we can
calculate the membrane potential of the Spike Response Model or the
integrate-and-fire neuron from Eq. (5.1) or (5.2),
respectively. In the deterministic model the next spike occurs when u
reaches the threshold
of the last spike, we can
calculate the membrane potential of the Spike Response Model or the
integrate-and-fire neuron from Eq. (5.1) or (5.2),
respectively. In the deterministic model the next spike occurs when u
reaches the threshold ![]() . In order to introduce some variability
into the neuronal spike generator, we replace the strict threshold by a
stochastic firing criterion. In the noisy threshold model, spikes can occur
at any time with a probability density,
. In order to introduce some variability
into the neuronal spike generator, we replace the strict threshold by a
stochastic firing criterion. In the noisy threshold model, spikes can occur
at any time with a probability density,
Since u on the right-hand side of Eq. (5.41) is a function of time,
the firing intensity ![]() is time-dependent as well. In view of Eqs. (5.1)
and (5.2), we write
is time-dependent as well. In view of Eqs. (5.1)
and (5.2), we write
![\centerline{\includegraphics[width=60mm]{noise-escape-mod.eps}}](img737.gif) |
Is Eq. (5.42) a sufficiently general noise model? We have seen in
Chapter 2.2 that the concept of a pure voltage threshold is
questionable. More generally, the spike trigger process could, for example,
also depend on the slope
![]() = du/dt with which the `threshold' is
approached. In the noisy threshold model, we may therefore also consider an
escape rate (or hazard) which depends not only on u but also on its
derivative
= du/dt with which the `threshold' is
approached. In the noisy threshold model, we may therefore also consider an
escape rate (or hazard) which depends not only on u but also on its
derivative ![]()
Note that the hazard ![]() in Eq. (5.43) is implicitly
time-dependent, via the membrane potential
u(t|
in Eq. (5.43) is implicitly
time-dependent, via the membrane potential
u(t|![]() ). In an even more
general model, we could in addition include an explicit time dependence,
e.g., to account for a reduced spiking probability immediately after the spike
at
). In an even more
general model, we could in addition include an explicit time dependence,
e.g., to account for a reduced spiking probability immediately after the spike
at ![]() . In the following examples we will stick to the hazard function as
defined by Eq. (5.42).
. In the following examples we will stick to the hazard function as
defined by Eq. (5.42).
We have motivated the escape model by a noisy version of the threshold process. In order to explore the relation between noisy and deterministic threshold models, we consider an escape function f defined as
![\centerline{\includegraphics[height=35mm,width=50mm]{esc-step.eps}}](img740.gif) |
How can we `soften' the sharp threshold? A simple choice for a soft threshold is an exponential dependence,
Finally, we can also use a sigmoidal escape rate (Wilson and Cowan, 1972; Abeles, 1982),
We want to motivate the sigmoidal escape rate by a model with stochastic
threshold in discrete time
tn = n ![]() . After each time step
of length
. After each time step
of length ![]() , a new value of the threshold is chosen from a Gaussian
distribution of threshold values
, a new value of the threshold is chosen from a Gaussian
distribution of threshold values
![]() with mean
with mean ![]() ,
,
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=35mm,width...
...f B}
\par\includegraphics[height=35mm,width=60mm]{gauss-erf.eps}
\end{minipage}](img770.gif) |
Instead of a model with stochastic threshold, we can also consider a model
fixed threshold, but a membrane potential
u(tn) + ![]() u(tn) with a stochastic component
u(tn) with a stochastic component ![]() u. If
u. If ![]() u is chosen at
each time step independently from a Gaussian distribution with variance
u is chosen at
each time step independently from a Gaussian distribution with variance
![]() and vanishing mean, we arrive again at formula
(5.47) (Abeles, 1982; Geisler and Goldberg, 1966; Weiss, 1966).
and vanishing mean, we arrive again at formula
(5.47) (Abeles, 1982; Geisler and Goldberg, 1966; Weiss, 1966).
The sigmoidal escape rate (5.51) has been motivated here for models
in discrete time. There are two potential problems. First, if we keep
![]() fixed and take
fixed and take
![]()
![]() 0 we do not recover the
deterministic threshold model.
Thus the low-noise limit is problematic. Second, since the
firing intensity diverges for
0 we do not recover the
deterministic threshold model.
Thus the low-noise limit is problematic. Second, since the
firing intensity diverges for
![]()
![]() 0, simulations
will necessarily depend on
the discretization
0, simulations
will necessarily depend on
the discretization ![]() .
This is due to the fact that the bandwidth of the
noise is limited by
.
This is due to the fact that the bandwidth of the
noise is limited by
![]() because a new value of
because a new value of
![]() or
or
![]() u is chosen at intervals
u is chosen at intervals ![]() . For
. For
![]()
![]() 0, the bandwidth
and hence the noise power diverge. Despite its problems, the sigmoidal escape
rate is also used in neuronal models in continuous time and either motivated
by a Gaussian distribution of threshold values (Wilson and Cowan, 1972) or else for
fixed threshold
0, the bandwidth
and hence the noise power diverge. Despite its problems, the sigmoidal escape
rate is also used in neuronal models in continuous time and either motivated
by a Gaussian distribution of threshold values (Wilson and Cowan, 1972) or else for
fixed threshold ![]() by a Gaussian distribution of membrane potentials
with band-limited noise (Abeles, 1982; Weiss, 1966). If we use
Eq. (5.47) in continuous time, the time scale
by a Gaussian distribution of membrane potentials
with band-limited noise (Abeles, 1982; Weiss, 1966). If we use
Eq. (5.47) in continuous time, the time scale ![]() becomes
a free parameter and should be taken proportional to the correlation time
becomes
a free parameter and should be taken proportional to the correlation time
![]() of the noise
of the noise ![]() u in the membrane potential (i.e.,
proportional to the inverse of the noise bandwidth). If the correlation time
is short, the model becomes closely related to continuous-time escape rate
models (Weiss, 1966). A `natural' correlation time of the membrane potential
will be calculated in Section 5.5 in the context of
stochastic spike arrival.
u in the membrane potential (i.e.,
proportional to the inverse of the noise bandwidth). If the correlation time
is short, the model becomes closely related to continuous-time escape rate
models (Weiss, 1966). A `natural' correlation time of the membrane potential
will be calculated in Section 5.5 in the context of
stochastic spike arrival.
In the previous example, we have started from a model in discrete time and
found that the limit of continuous time is not without problems. Here we want
to start from a model in continuous time and discretize time as it is often
done in simulations. In a straightforward discretization scheme, we
calculate the probability of firing during a time step ![]() t of a neuron
that has fired the last time at
t of a neuron
that has fired the last time at ![]() as
as
![]()
![]() (t'|
(t'|![]() ) dt'
) dt' ![]()
![]() (t|
(t|![]() )
) ![]() t. For
u
t. For
u ![]()
![]() , the hazard
, the hazard
![]() (t|
(t|![]() ) = f[u(t|
) = f[u(t|![]() ) -
) - ![]() ] can take large values; see,
e.g., Eq. (5.45). Thus
] can take large values; see,
e.g., Eq. (5.45). Thus ![]() t must be taken extremely short so
as to guarantee
t must be taken extremely short so
as to guarantee
![]() (t|
(t|![]() )
) ![]() t < 1.
t < 1.
In order to arrive at an improved discretization scheme, we calculate the
probability that a neuron does not fire in a time step ![]() t.
Since the integration of Eq. (5.6) over a finite time
t.
Since the integration of Eq. (5.6) over a finite time ![]() t yields an exponential factor analogous to Eq. (5.7), we arrive
at a firing probability
t yields an exponential factor analogous to Eq. (5.7), we arrive
at a firing probability
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=35mm,width...
...r\includegraphics[height=35mm,width=60mm]{discrete-srm-beta.eps}
\end{minipage}](img777.gif) |
In this section, we combine the escape rate model with the concepts of
renewal theory and calculate the input-dependent interval distribution
PI(t|![]() ) for escape rate models.
) for escape rate models.
We recall Eq. (5.9) and express the interval distribution in terms
of the hazard ![]() ,
,
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=35mm,width...
...r\includegraphics[height=35mm,width=60mm]{pout.dat.gain-srm.eps}
\end{minipage}](img778.gif) |
To do so, we express ![]() by the escape rate. In order to keep the
notation simple, we suppose that the escape rate f is a function of u
only. We insert Eq. (5.42) into Eq. (5.53) and obtain
by the escape rate. In order to keep the
notation simple, we suppose that the escape rate f is a function of u
only. We insert Eq. (5.42) into Eq. (5.53) and obtain
We study the model SRM0 defined in Eq. (5.55) for absolute refractoriness
In this example we show that interval distributions are particularly simple if
a linear escape rate is adopted. We start with the non-leaky
integrate-and-fire model. In the limit of
![]()
![]()
![]() , the membrane
potential of an integrate-and-fire neuron is
, the membrane
potential of an integrate-and-fire neuron is
For a leaky integrate-and-fire neuron with constant input I0, the membrane potential is
| u(t| |
(5.61) |
| (5.62) |
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=35mm,width...
...ludegraphics[height=35mm,width=60mm]{pout.dat.potential-per.eps}
\end{minipage}](img795.gif) |
We study the model SRM0 as defined in Eq. (5.55) with periodic
input
I(t) = I0 + I1cos(![]() t). This leads to an input
potential
h(t) = h0 + h1 cos(
t). This leads to an input
potential
h(t) = h0 + h1 cos(![]() t +
t + ![]() ) with bias h0 = I0 and a periodic component with a certain amplitude h1 and phase
) with bias h0 = I0 and a periodic component with a certain amplitude h1 and phase
![]() . We choose a refractory kernel with absolute and relative
refractoriness defined as
. We choose a refractory kernel with absolute and relative
refractoriness defined as
Suppose that a spike has occurred at ![]() = 0. The probability
density that the next spike occurs at time t is given by
PI(t|
= 0. The probability
density that the next spike occurs at time t is given by
PI(t|![]() )
and can be calculated from Eq. (5.53). The result is shown in
Fig. 5.10. We note that the periodic component of the input is
well represented in the response of the neuron. This example illustrates how
neurons in the auditory system can transmit stimuli of frequencies higher
than the mean firing rate of the neuron; see Chapter 12.5. We
emphasize that the threshold in Fig. 5.10 is at
)
and can be calculated from Eq. (5.53). The result is shown in
Fig. 5.10. We note that the periodic component of the input is
well represented in the response of the neuron. This example illustrates how
neurons in the auditory system can transmit stimuli of frequencies higher
than the mean firing rate of the neuron; see Chapter 12.5. We
emphasize that the threshold in Fig. 5.10 is at ![]() =1.
Without noise there would be no output spike. On the other hand, at very high
noise levels, the modulation of the interval distribution would be much
weaker. Thus a certain amount of noise is beneficial for signal
transmission. The existence of a optimal noise level
is a phenomenon called stochastic resonance and will be discussed below in
Section 5.8.
=1.
Without noise there would be no output spike. On the other hand, at very high
noise levels, the modulation of the interval distribution would be much
weaker. Thus a certain amount of noise is beneficial for signal
transmission. The existence of a optimal noise level
is a phenomenon called stochastic resonance and will be discussed below in
Section 5.8.
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
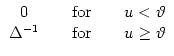
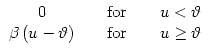
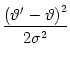
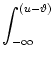
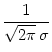 exp
exp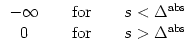 .
.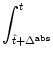 r(t')dt'
r(t')dt'