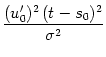One of the aims of noisy neuron models is to mimic the large variability of interspike intervals found, e.g., in vertebrate cortex. To arrive at broad interval distributions, it is not just sufficient to introduce noise into a neuron model. Apart from the noise level, other neuronal parameters such as the firing threshold or a bias current have to be tuned so as to make the neuron sensitive to noise. In this section we introduce a distinction between super- and subthreshold stimulation (Bugmann et al., 1997; König et al., 1996; Shadlen and Newsome, 1994; Troyer and Miller, 1997; Abeles, 1991). In Section 5.7 we will show that, in the subthreshold regime, there is a close relation between the two different noise models discussed above, viz., escape noise (cf. Section 5.3) and diffusive noise (cf. Section 5.5). Finally, in Section 5.8 we turn to the phenomenon of stochastic resonance and discuss signal transmission in the subthreshold regime.
An arbitrary time-dependent stimulus I(t) is called subthreshold, if it generates a membrane potential that stays - in the absence of noise - below the firing threshold. Due to noise, however, even subthreshold stimuli can induce action potentials. Stimuli that induce spikes even in a noise-free neuron are called superthreshold.
The distinction between sub- and superthreshold stimuli has important consequences for the firing behavior of neurons in the presence of noise. To see why, let us consider an integrate-and-fire neuron with constant input I0 for t > 0. Starting from u(t = 0) = ur, the trajectory of the membrane potential is
![\centerline{\includegraphics[width=60mm]{Fig-Gauss2a.eps}}](img902.gif) |
We now add diffusive noise. In the superthreshold regime, noise has little influence, except that it broadens the interspike interval distribution. Thus, in the superthreshold regime, the spike train in the presence of diffusive noise, is simply a noisy version of the regular spike train of the noise-free neuron.
On the other hand, in the subthreshold regime, the spike train changes qualitatively, if noise is switched on; see (König et al., 1996) for a review. Stochastic background input turns the quiescent neuron into a spiking one. In the subthreshold regime, spikes are generated by the fluctuations of the membrane potential, rather than by its mean (Bugmann et al., 1997; Feng, 2001; Shadlen and Newsome, 1994; Troyer and Miller, 1997; Abeles, 1991). The interspike interval distribution is therefore broad; see Fig. 5.20.
![\begin{minipage}{0.45\textwidth}
{\bf A}
\par\includegraphics[height=35mm,width...
...r\includegraphics[height=35mm,width=60mm]{pout.dat.PSTH-0.8.eps}
\end{minipage}](img903.gif) |
For small noise amplitude
0 < ![]()
![]() u
u![]() -
- ![]() , the interval
distribution is centered at s0. Its width can be estimated from the width
of the fluctuations
, the interval
distribution is centered at s0. Its width can be estimated from the width
of the fluctuations
![]()
![]() u2
u2![]()
![]() of the free
membrane potential; cf. Eq. (5.99). Since the membrane potential
crosses the threshold with slope u0', there is a scaling factor
u0' = du0(t)/dt evaluated at t = s0; cf. Fig. 5.19. The
interval distribution is therefore approximately given by a Gaussian with mean
s0 and width
of the free
membrane potential; cf. Eq. (5.99). Since the membrane potential
crosses the threshold with slope u0', there is a scaling factor
u0' = du0(t)/dt evaluated at t = s0; cf. Fig. 5.19. The
interval distribution is therefore approximately given by a Gaussian with mean
s0 and width
![]() /
/![]() u0' (Tuckwell, 1988), i.e.,
u0' (Tuckwell, 1988), i.e.,
Figures 5.20e and 5.20f show that interval distributions in the super- and subthreshold regime look quite differently. To quantify the width of the interval distribution, neuroscientists often evaluate the coefficient of variation, short CV, defined as the ratio of the variance and the mean squared,
We study a Poisson neuron with absolute refractory period
![]() .
For
t -
.
For
t - ![]() >
> ![]() , the neuron is supposed to fire
stochastically with rate r. The interval distribution is given in
Eq. (5.20) with mean
, the neuron is supposed to fire
stochastically with rate r. The interval distribution is given in
Eq. (5.20) with mean
![]() s
s![]() =
= ![]() + 1/r
and variance
+ 1/r
and variance
![]()
![]() s2
s2![]() = 1/r2. The coefficient of
variation is therefore
= 1/r2. The coefficient of
variation is therefore
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
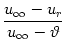 .
.