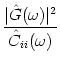Our considerations regarding step current input can be
generalized to an arbitrary input current I(t)
that is fluctuating around a mean value of I0.
We study a population of independent integrate-and-fire or SRM0 neurons.
The input current
I(t) = I0 + ![]() I(t) generates an
input potential
I(t) generates an
input potential
| h(t) = |
(7.51) |
| I(t) = I0 + I1 cos( |
(7.52) |
We assume that the population is close to a state of asynchronous firing,
viz.,
A(t) = A0 + ![]() A(t). The linear response of the population to
the change in the input potential h is given by Eq. (7.3) which
can be solved for
A(t). The linear response of the population to
the change in the input potential h is given by Eq. (7.3) which
can be solved for ![]() A by taking the Fourier transform.
For
A by taking the Fourier transform.
For
![]()
![]() 0 we find
0 we find
The function
![]() (
(![]() ), defined by Eq. (7.57), describes
the (linear) response
), defined by Eq. (7.57), describes
the (linear) response
![]() (
(![]() ) of a population of spiking neurons to
a periodic signal
) of a population of spiking neurons to
a periodic signal
![]() (
(![]() ). It is also called the
(frequency-dependent) gain of the system. Inverse Fourier transform of
Eq. (7.57) yields
). It is also called the
(frequency-dependent) gain of the system. Inverse Fourier transform of
Eq. (7.57) yields
We can compare the amplitude of an input current at frequency ![]() with
the amplitude of the response. The ratio
with
the amplitude of the response. The ratio
| (7.56) |
In the following examples signal we will study transmission properties of a population of neurons with different noise models. In particular, we will see that for slow noise in the parameters (e.g. noise in the reset) signal transmission is possible at very high frequencies (that is, there is no cut-off frequency) (Gerstner, 2000b; Knight, 1972a). On the other hand, for escape noise models the cut-off frequency depends on the noise level. For a large amount of escape noise, the cut-off frequency is given by the the inverse of the membrane time constant (Gerstner, 2000b). Finally, diffusive noise models have a cut-off frequency if the noise input is white (standard diffusion model), but do not have a cut-off frequency if the noise has a long correlation time (Brunel et al., 2001).
Even if there is no cut-off frequency for the transmission of fast input currents, we may not conclude that real neurons are infinitely fast. In fact, a finite time constant of synaptic channels leads to a frequency cut-off for the input current which may enter the cell. In this sense, it is the time constant of the synaptic current which determines the cut-off frequency of the population. The membrane time constant is of minor influence (Gerstner, 2000b; Treves, 1993; Knight, 1972a).
![\vbox{
\vspace{5mm}
\hbox{
{\bf A}
\hspace{70mm}
{\bf B}
}
\par\hbox{
\hs...
...ight=20mm,width=40mm]{Figs-ch-signal/pout.dat.signal-A.eps}
}
\vspace{2mm}
}](img1238.gif) |
In this example, we consider integrate-and-fire neurons with noisy reset; cf.
Chapter 5.4. For noisy reset the interval distribution
in the stationary state is a Gaussian
P0(s) = ![]()
![]() (s - T0)
with mean T0 and width
(s - T0)
with mean T0 and width ![]() ;
cf. Eq. (7.38). Fourier transform of the interval
distribution yields
;
cf. Eq. (7.38). Fourier transform of the interval
distribution yields
| (7.57) |
| (7.58) |
We adjust the bias current I0
so that the mean
interspike interval of the neurons is T0 = 8ms.
In Fig. 7.9A we have plotted the gain
|![]() (
(![]() )| = |
)| = |![]() (
(![]() )/
)/![]() (
(![]() )| as a function of the
stimulation frequency
f =
)| as a function of the
stimulation frequency
f = ![]() /(2
/(2![]() ). For a medium noise level of
). For a medium noise level of ![]() = 2ms, the signal gain
has a single resonance at
f = 1/T0 = 125Hz. For lower noise, further
resonances at multiples of 125 Hz appear. For a variant of the noisy reset
model, a result closely related to Eq. (7.57) has been derived by
Knight (1972a).
= 2ms, the signal gain
has a single resonance at
f = 1/T0 = 125Hz. For lower noise, further
resonances at multiples of 125 Hz appear. For a variant of the noisy reset
model, a result closely related to Eq. (7.57) has been derived by
Knight (1972a).
Independently of the noise level, we obtain for integrate-and-fire neurons for
![]()
![]() 0 the result
|
0 the result
|![]() (0)| = JextA0[1 - exp(- T0/
(0)| = JextA0[1 - exp(- T0/![]() )]/(u' T0). Most interesting is the behavior in the
high-frequency limit. For
)]/(u' T0). Most interesting is the behavior in the
high-frequency limit. For
![]()
![]()
![]() we find
|
we find
|![]() (
(![]() )|
)|![]() RA0/(u'
RA0/(u' ![]() ), hence
), hence
So far we have discussed results of the linearized theory; viz., Eqs.
(7.41) and (7.57). The behavior of the full non-linear
system is shown in Fig. 7.10. A population of unconnected SRM0
neurons is stimulated by a time-dependent input current which was generated as
a superposition of 4 sinusoidal components with frequencies at 9, 47, 111 and
1000Hz which have been chosen arbitrarily. The activity equation
A(t) = ![]() PI(t|
PI(t|![]() ) A(
) A(![]() ) d
) d![]() been integrated with time steps of 0.05ms and the
results are compared with those of a simulation of a population of 4000
neurons. The 1kHz component of the signal I(t) is clearly reflected in
the population A(t). Theory and simulation are in excellent agreement.
been integrated with time steps of 0.05ms and the
results are compared with those of a simulation of a population of 4000
neurons. The 1kHz component of the signal I(t) is clearly reflected in
the population A(t). Theory and simulation are in excellent agreement.
![\vbox{
\vspace{5mm}
\par\hbox{
\hspace{30mm}
\par\includegraphics[height=40mm,width=60mm]{Figs-ch-signal/pout.dat.signal-aoft-all.ps}
}
\vspace{2mm}
}](img1255.gif) |
We have seen in the preceding section, that noisy reset is rather exceptional
in the sense that the transient remains sharp even in the limit of high
noise. To study the relevance of the noise model, we return to
Eq. (7.57). The signal gain
![]() (
(![]() ) = | A(
) = | A(![]() )/I(
)/I(![]() )| is proportional to
)| is proportional to
![]() (
(![]() ). If the kernel
). If the kernel
![]() (x) is broad, its Fourier transform
(x) is broad, its Fourier transform
![]() (
(![]() ) will fall off to
zero at high frequencies and so does the signal gain
) will fall off to
zero at high frequencies and so does the signal gain
![]() (
(![]() ). In
Fig. 7.9B we have plotted the signal gain
). In
Fig. 7.9B we have plotted the signal gain
![]() (
(![]() ) for
integrate-and-fire neurons with escape noise at different noise levels. At
low noise, the result for escape noise is similar to that of reset noise
(compare Figs. 7.9A and B) except for a drop of the gain at high
frequencies. Increasing the noise level, however, lowers the signal gain of
the system. For high noise (long-dashed line in Fig. 7.9B the
signal gain at 1000 Hz is ten times lower than the gain at zero frequency.
The cut-off
frequency depends on the noise level. Note that for escape noise, the gain at
zero frequency also changes with the level of noise.
) for
integrate-and-fire neurons with escape noise at different noise levels. At
low noise, the result for escape noise is similar to that of reset noise
(compare Figs. 7.9A and B) except for a drop of the gain at high
frequencies. Increasing the noise level, however, lowers the signal gain of
the system. For high noise (long-dashed line in Fig. 7.9B the
signal gain at 1000 Hz is ten times lower than the gain at zero frequency.
The cut-off
frequency depends on the noise level. Note that for escape noise, the gain at
zero frequency also changes with the level of noise.
It is possible to calculate the signal transmission properties of
integrate-and-fire neurons with diffusive noise by a linearization of the
population density equation (6.21) about the stationary
membrane potential distribution p0(u). The resulting formula
(Brunel et al., 2001) is rather complicated but can be evaluated numerically.
It is found that in the standard diffusion
model the gain
|![]() (
(![]() )|
decays as
1/
)|
decays as
1/![]() for large
for large ![]() .
Thus the gain exhibits a cut-off
frequency similar to that found in the escape noise model.
.
Thus the gain exhibits a cut-off
frequency similar to that found in the escape noise model.
Standard diffusive noise corresponds to a drive by stochastic ![]() current
pulses, which is usually motivated as a description of stochastic spike
arrival; cf. Chapter 5. In a more realistic model of stochastic
spike arrival, input spikes evoke a current pulse of finite width.
The duration of the current pulse is characterized by the synaptic time
constant
current
pulses, which is usually motivated as a description of stochastic spike
arrival; cf. Chapter 5. In a more realistic model of stochastic
spike arrival, input spikes evoke a current pulse of finite width.
The duration of the current pulse is characterized by the synaptic time
constant ![]() . In that case, the effective noisy input current has
correlations on the time scale of
. In that case, the effective noisy input current has
correlations on the time scale of ![]() . If
. If
![]() > 1/A0, the noise is
`slow' compared to the intrinsic firing rate of the neuron. It is found that
with such a slow noise, the
> 1/A0, the noise is
`slow' compared to the intrinsic firing rate of the neuron. It is found that
with such a slow noise, the
![]() (
(![]() ) has no cut-off frequency
(Brunel et al., 2001). In this limit, the gain factor is therefore similar to that
of the stochastic reset model. In other words, we have the generic result
that for `fast' noise the gain factor has a cut-off frequency whereas for
`slow' noise it has not.
) has no cut-off frequency
(Brunel et al., 2001). In this limit, the gain factor is therefore similar to that
of the stochastic reset model. In other words, we have the generic result
that for `fast' noise the gain factor has a cut-off frequency whereas for
`slow' noise it has not.
So far we have considered the signal transmission properties of a large
population in the limit
N![]()
![]() . In this case the population activity
can be considered as a continuous signal, even though individual neurons emit
short pulse-like action potentials. For a finite number N of neurons,
however, the population activity A(t) will fluctuate around a time-dependent
mean. In this section we want to estimate the amplitude of the fluctuations.
. In this case the population activity
can be considered as a continuous signal, even though individual neurons emit
short pulse-like action potentials. For a finite number N of neurons,
however, the population activity A(t) will fluctuate around a time-dependent
mean. In this section we want to estimate the amplitude of the fluctuations.
For independent neurons that are stimulated by a constant current I0, we can calculate the noise spectrum of the population activity using the methods discussed in Chapter 5. In fact, the noise spectrum CAA of the population activity is proportional to the Fourier transform of the autocorrelation function of a single-neuron:
| CAA( |
(7.60) |
If the amplitude of the periodic stimulation is small, the noise term of the
population activity can be estimated from the stationary
autocorrelation function. The signal-to-noise ratio at frequency ![]() is
is
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
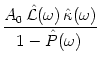
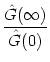
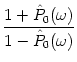
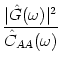 = N
= N