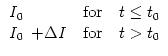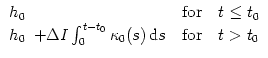How quickly can a population of neurons respond to a rapid change in the
input? We know from reaction time experiments that the response of humans and
animals to new stimuli can be very fast (Thorpe et al., 1996). We therefore expect
that the elementary processing units, i.e., neurons or neuronal populations
should also show a rapid response. In this section we concentrate on one
element of the problem of rapid reaction time and study the response of the
population activity to a rapid change in the input. To keep the arguments as
simple as possible, we consider an input which has a constant value I0
for t < t0 and changes then abruptly to a new value
I0 + ![]() I.
Thus
I.
Thus
For the sake of simplicity, we consider a population of independent
integrate-and-fire or SRM0 neurons without lateral coupling. Given the
current
Iext(t), the input potential can be determined from
h(t) = ![]()
![]() (s) Iext(t - s) ds. For t
(s) Iext(t - s) ds. For t![]() t0,
the input potential has then a value
h0 = R I0 where we have used
t0,
the input potential has then a value
h0 = R I0 where we have used
![]()
![]() (s)ds = R. For t > t0, the input potential h changes due to
the additional current
(s)ds = R. For t > t0, the input potential h changes due to
the additional current ![]() I so that
I so that
Let us suppose that for t < t0 the network is in a state of asynchronous firing so that the population activity is constant,
A(t) = A0 for t![]() t0; cf. Chapter 6.4. As soon as
the input is switched on at time t = t0, the population activity will change
t0; cf. Chapter 6.4. As soon as
the input is switched on at time t = t0, the population activity will change
| A(t) = A0 + |
(7.40) |
![\hbox{{\bf A}
\hspace{60mm}
{\bf B}}
\par\hbox{
\hspace{10mm}
\includegraphi...
...cs[height=15mm,width=40mm]{Figs-ch-signal/pout.dat.ioft.ps}
}
\par\vspace{5mm}](img1214.gif) |
In the noiseless case, neurons which receive a constant input I0 fire regularly with some period T0. For t < t0, the mean activity is simply A0 = 1/T0. The reason is that, for a constant activity, averaging over time and averaging over the population are equivalent; cf. Chapter 6.4.
Let us consider a neuron which has fired exactly at t0. Its next spike
occurs at t0 + T where T is given by the threshold condition
ui(t0 + T) = ![]() . We focus on the initial phase of the transient and apply the
noise-free kernel
. We focus on the initial phase of the transient and apply the
noise-free kernel
![]() (x)
(x) ![]()
![]() (x);
cf. Tab. 7.1.
If we insert the
(x);
cf. Tab. 7.1.
If we insert the ![]() function into Eq. (7.46)
we find
function into Eq. (7.46)
we find
| (7.43) |
In this example we apply Eq. (7.48) to integrate-and-fire neurons. The response kernel is
A similar result holds for a population of SRM0 neurons.
The initial transient of
SRM0 is identical to that of integrate-and-fire neurons;
cf. Fig. 7.5.
A subtle difference, however, occurs during the late
phase of the transient.
For
integrate-and-fire neurons the transient is over as soon as each neuron has
fired once.
After the next reset, all neurons fire periodically
with a new period T that corresponds to
the constant input
I0 + ![]() I.
A population of SRM0 neurons, however, reaches a periodic state
only asymptotically.
The reason is that the interspike interval T of
SRM0 neurons [which is given by the threshold condition
h(t) =
I.
A population of SRM0 neurons, however, reaches a periodic state
only asymptotically.
The reason is that the interspike interval T of
SRM0 neurons [which is given by the threshold condition
h(t) = ![]() -
- ![]() (T)] depends on the momentary
value of the input potential h(t).
(T)] depends on the momentary
value of the input potential h(t).
![\vbox{
\vspace{0mm}
\hbox{
{\bf A}
\hspace{60mm}
{\bf B}
}
\par\hbox{
\hs...
...ht=30mm,width=55mm]{Figs-ch-signal/pout.WC-Abaroft.eps}
\par }
\vspace{0mm}
}](img1222.gif) |
So far, we have considered noiseless neurons. We have seen that after an
initial sharp transient the population activity approaches a new periodic
state where the activity oscillates with period T. In the presence of
noise, we expect that the network approaches - after a transient - a new
asynchronous state with stationary activity
A0 = g(I0 + ![]() I);
cf. Chapter 6.4.
I);
cf. Chapter 6.4.
In Fig. 7.6A illustrates the response of a population of noisy neurons to a step current input. The population activity responds instantaneously as soon as the additional input is switched on. Can we understand the sharply peaked transient? Before the abrupt change the input was stationary and the population in a state of asynchronous firing. Asynchronous firing was defined as a state with constant activity so that at any point in time some of the neurons fire, others are in the refractory period, again others approach the threshold. There is always a group of neuron whose potential is just below threshold. An increase in the input causes those neurons to fire immediately - and this accounts for the strong population response during the initial phase of the transient.
As we will see in the example below, the above consideration is strictly valid only for neurons with slow noise in the parameters, e.g., noisy reset as introduced in Chapter 5.4. In models based on the Wilson-Cowan differential equation the transient does not exhibit such a sharp initial peak; cf. Fig. 7.6B.
For diffusive noise models the picture is more complicated. A rapid response occurs if the current step is sufficiently large and the noise level not too high. On the other hand, for high noise and small current steps the response is slow. The question of whether neuronal populations react rapidly or slowly depends therefore on many aspects, in particular on the type of noise and the type of stimulation. It can be shown that for diffusive noise that is low-pass filtered by a slow synaptic time constant (i.e., cut-off frequency of the noise lower than the neuronal firing rate) the response is sharp, independent of the noise amplitude. On the other hand, for white noise the response depends on the noise amplitude and the membrane time constant (Brunel et al., 2001).
For a mathematical discussion of the transient behavior, it is sufficient to
consider the equation that describes the initial phase of the linear response
to a sudden onset of the input potential; cf. Eq. (7.46).
Table 7.1 summarizes the kernel
![]() (x) that is at the heart of
Eq. (7.46) for several noise models. In the limit of low noise,
the choice of noise model is irrelevant - the transient response is
proportional to the derivative of the potential,
(x) that is at the heart of
Eq. (7.46) for several noise models. In the limit of low noise,
the choice of noise model is irrelevant - the transient response is
proportional to the derivative of the potential,
![]() A
A ![]() h'. If the level of noise is increased, a population of neurons with slow
noise (e.g., with noisy reset) retains its sharp transients since the kernel
h'. If the level of noise is increased, a population of neurons with slow
noise (e.g., with noisy reset) retains its sharp transients since the kernel
![]() is proportional to h',
is proportional to h',
Neurons with escape noise turn in the high-noise limit to a different regime
where the transients follow h rather than h'. To see why, we recall that
the kernel
![]() essentially describes a low-pass filter;
cf. Fig. 7.3.
The time constant
of the filter increases with the noise level and hence the response switches
from a behavior proportional to h' to a behavior proportional to h.
essentially describes a low-pass filter;
cf. Fig. 7.3.
The time constant
of the filter increases with the noise level and hence the response switches
from a behavior proportional to h' to a behavior proportional to h.
The width of the kernel
![]() (x)
in Eq. (7.46)
depends on the noise level.
For low noise, the kernel is
sharply peaked at x = 0 and
can be approximated by a Dirac
(x)
in Eq. (7.46)
depends on the noise level.
For low noise, the kernel is
sharply peaked at x = 0 and
can be approximated by a Dirac ![]() function.
The response
function.
The response ![]() A
of the population activity is sharp
since it is proportional to the derivative
of the input potential.
A
of the population activity is sharp
since it is proportional to the derivative
of the input potential.
For high noise, the kernel is broad and the response becomes proportional to the input potential; cf. Fig. 7.7.
![\vbox{
\vspace{0mm}
\hbox{
{\bf A}
\hspace{60mm}
{\bf B}
}
\par\hbox{
\hspace{5m...
...ght=30mm,width=55mm]{Figs-ch-signal/Aoft-A-slowtrans.eps}
\par }
\vspace{0mm}
}](img1223.gif) |
In Chapter 6.3, we have introduced the Wilson-Cowan
differential equations which are summarized here for a population of
independent neurons,
| h(t) = h0 + R |
(7.49) |
For neurons with noisy reset, the kernel
![]() is a Dirac
is a Dirac ![]() function;
cf. Tab. 7.1. As in the noiseless case, the initial transient
is therefore proportional to the derivative of h. After this initial phase
the reset noise leads to a smoothing of subsequent oscillations so that the
population activity approaches rapidly a new asynchronous state;
cf. Fig. 7.6A. The initial transient, however, is sharp.
function;
cf. Tab. 7.1. As in the noiseless case, the initial transient
is therefore proportional to the derivative of h. After this initial phase
the reset noise leads to a smoothing of subsequent oscillations so that the
population activity approaches rapidly a new asynchronous state;
cf. Fig. 7.6A. The initial transient, however, is sharp.
In this example, we present qualitative arguments to show that, in the limit
of low noise, a population of spiking neurons with diffusive noise will
exhibit an immediate response to a strong step current input. We have
seen in the noise-free case, that the rapid response is due the derivative
h' in the compression factor. In order to understand, why the derivative of
h comes into play, let us consider, for the moment, a finite step in the
input potential
h(t) = h0 + ![]() h
h ![]() (t - t0). All neurons i
which are hovering below threshold so that their potential ui(t0) is
between
(t - t0). All neurons i
which are hovering below threshold so that their potential ui(t0) is
between
![]() -
- ![]() h and
h and ![]() will be put above threshold and
fire synchronously at t0. Thus, a step in the potential causes a
will be put above threshold and
fire synchronously at t0. Thus, a step in the potential causes a
![]() -pulse in the activity
-pulse in the activity
![]() A(t)
A(t) ![]()
![]() (t - t0)
(t - t0) ![]() h'(t0). In Fig. 7.8a we have used a current step
(7.42) [the same step current as in Fig. 7.5]. The
response at low noise (top) has roughly the form
h'(t0). In Fig. 7.8a we have used a current step
(7.42) [the same step current as in Fig. 7.5]. The
response at low noise (top) has roughly the form
![]() A(t)
A(t) ![]() h'(t)
h'(t) ![]()
![]() (t - t0) as expected. The rapid transient is slightly less
pronounced than for noisy reset, but nevertheless clearly visible; compare
Figs. 7.6A and 7.8A. As the amplitude of the noise
grows, the transient becomes less sharp. Thus there is a transition from a
regime where the transient is proportional to h' (Fig. 7.8A) to
another regime where the transient is proportional to h
(Fig. 7.8B). What are the reasons for the change of behavior?
(t - t0) as expected. The rapid transient is slightly less
pronounced than for noisy reset, but nevertheless clearly visible; compare
Figs. 7.6A and 7.8A. As the amplitude of the noise
grows, the transient becomes less sharp. Thus there is a transition from a
regime where the transient is proportional to h' (Fig. 7.8A) to
another regime where the transient is proportional to h
(Fig. 7.8B). What are the reasons for the change of behavior?
The simple argument from above based
on a potential step
![]() h > 0 only holds
for a finite step size which is
at least of the order of the noise amplitude
h > 0 only holds
for a finite step size which is
at least of the order of the noise amplitude ![]() .
With diffusive noise, the threshold acts as an absorbing
boundary. Therefore the density of neurons with potential
ui vanishes for
ui
.
With diffusive noise, the threshold acts as an absorbing
boundary. Therefore the density of neurons with potential
ui vanishes for
ui![]()
![]() ;
cf. Chapter 6.2.
Thence, for
;
cf. Chapter 6.2.
Thence, for
![]() h
h![]() 0 the proportion of neurons
which are instantaneously put across threshold is 0.
In a stationary state, the 'boundary layer' with low density
is of the order
0 the proportion of neurons
which are instantaneously put across threshold is 0.
In a stationary state, the 'boundary layer' with low density
is of the order ![]() ; e.g.,
cf. Eq. (6.28).
A potential step
; e.g.,
cf. Eq. (6.28).
A potential step
![]() h >
h > ![]() puts a significant
proportion of neurons above threshold and leads to a
puts a significant
proportion of neurons above threshold and leads to a
![]() -pulse in the activity. Thus the result that
the response is proportional to the derivative of the potential
is essentially valid in the low-noise regime.
-pulse in the activity. Thus the result that
the response is proportional to the derivative of the potential
is essentially valid in the low-noise regime.
![\vbox{
\vspace{0mm}
\hbox{
{\bf A}
\hspace{60mm}
{\bf B}
}
\par\hbox{
\hspace{5m...
...ght=30mm,width=55mm]{Figs-ch-signal/Aoft-C-smallstep.eps}
\par }
\vspace{0mm}
}](img1224.gif) |
On the other hand, we may also consider diffusive noise with large noise amplitude in the sub-threshold regime. In the limit of high noise, a step in the potential raises the instantaneous rate of the neurons, but does not force them to fire immediately. The response to a current step is therefore smooth and follows the potential h(t); cf. Fig. 7.8B. A comparison of Figs. 7.8 and 7.7 shows that the escape noise model exhibits a similar transition form sharp to smooth responses with increasing noise level. In fact, we have seen in Chapter 5 that diffusive noise can be well approximated by escape noise (Plesser and Gerstner, 2000). For the analysis of response properties with diffusive noise see Brunel et al. (2001).
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.