We have seen in the previous section that the state of asynchronous firing can loose stability towards certain oscillatory modes that are solutions of the linearized population equations. We are now going to investigate oscillatory modes in more detail and check whether a large-amplitude oscillation where all neurons are firing in ``lockstep'' can be a stable solution of the population equations.
We consider a homogeneous population of SRM0 or integrate-and-fire neurons
which is nearly perfectly synchronized and fires almost regularly with period
T. In order to analyze the existence and stability of a fully locked
synchronous oscillation we approximate the population activity by a sequence
of square pulses k,
k ![]() {0,±1,±2,...}, centered around
t = k T. Each pulse k has a certain half-width
{0,±1,±2,...}, centered around
t = k T. Each pulse k has a certain half-width ![]() and amplitude
(2
and amplitude
(2![]() )-1 - since all neurons are supposed to fire once in each
pulse. In order to check whether the fully synchronized state is a stable
solution of the population equation (6.75), we assume that the
population has already fired a couple of narrow pulses for t < 0 with widths
)-1 - since all neurons are supposed to fire once in each
pulse. In order to check whether the fully synchronized state is a stable
solution of the population equation (6.75), we assume that the
population has already fired a couple of narrow pulses for t < 0 with widths
![]()
![]() T, k
T, k![]() 0, and calculate the amplitude and width of
subsequent pulses. If we find that the amplitude of subsequent pulses
increases while their width decreases (i.e.,
0, and calculate the amplitude and width of
subsequent pulses. If we find that the amplitude of subsequent pulses
increases while their width decreases (i.e.,
![]()
![]() = 0), then we conclude that the fully locked state is stable.
= 0), then we conclude that the fully locked state is stable.
To make the above outline more explicit, we use
![\centerline{
\includegraphics[height=15mm,width=60mm]{Figs-ch-oscillations/Fig-rectangular.eps}
}](img1315.gif) |
As we will see below, the condition for stable locking of all neurons in the population can be stated as a condition on the slope of the input potential h at the moment of firing. More precisely, if the last population pulse occurred at about t = 0 with amplitude A(0) the amplitude of the population pulse at t = T increases, if h'(T) > 0:
The Locking Theorem is applicable for large populations that are already close to the fully synchronized state. A related but global locking argument has been presented by (Mirollo and Strogatz, 1990). The locking argument can be generalized to heterogeneous networks (Chow, 1998; Gerstner et al., 1993a) and to electrical coupling (Chow and Kopell, 2000). Synchronization in small networks has been discussed in, e.g., (Bose et al., 2000; Hansel et al., 1995; Chow, 1998; Ernst et al., 1995; van Vreeswijk, 1996; van Vreeswijk et al., 1994). For weak coupling, synchronization and locking can be systematically analyzed in the framework of phase models (Ermentrout and Kopell, 1984; Kopell, 1986; Kuramoto, 1975) or canonical neuron models (Izhikevich, 1999; Hansel et al., 1995; Ermentrout, 1996; Ermentrout et al., 2001; Hoppensteadt and Izhikevich, 1997).
Before we derive the locking condition for spiking neuron models, we illustrate the main idea by two examples.
In this example we will show that locking in a population of spiking neurons can be understood by simple geometrical arguments; there is no need to use the abstract mathematical framework of the population equations. It will turn out that the results are - of course - consistent with those derived from the population equation.
We study a homogeneous network of N identical neurons which are mutually
coupled with strength
wij = J0/N where J0 > 0 is a positive
constant. In other words, the (excitatory) interaction is scaled with one over
N so that the total input to a neuron i is of order one even if the number
of neurons is large (
N![]()
![]() ). Since we are interested in synchrony we
suppose that all neurons have fired simultaneously at
). Since we are interested in synchrony we
suppose that all neurons have fired simultaneously at
![]() = 0. When
will the neurons fire again?
= 0. When
will the neurons fire again?
Since all neurons are identical we expect that the next firing time will also be synchronous. Let us calculate the period T between one synchronous pulse and the next. We start from the firing condition of SRM0 neurons
![\begin{minipage}[t]{0.45\textwidth}
{\bf A}\\
\par\includegraphics[width=\text...
...cludegraphics[width=\textwidth]{Figs-ch-oscillations/lock9b.eps}
\end{minipage}](img1317.gif) |
What happens if synchrony at t = 0 was not perfect? Let us assume that one of
the neurons is slightly late compared to the others; Fig. 8.5B. It
will receive the input
J0 ![]() (t) from the others, thus the
right-hand side of (8.14) is the same. The left-hand side, however, is
different since the last firing was at
(t) from the others, thus the
right-hand side of (8.14) is the same. The left-hand side, however, is
different since the last firing was at ![]() instead of zero. The next
firing time is at
t = T +
instead of zero. The next
firing time is at
t = T + ![]() where
where ![]() is found from
is found from
We see from Fig. 8.5B that, in the case of excitatory coupling,
stable locking works nicely if the transmission delay
![]() is in
the range of the firing period, but slightly shorter so that firing occurs
during the rise time of the EPSP.
is in
the range of the firing period, but slightly shorter so that firing occurs
during the rise time of the EPSP.
Locking can also occur in networks with purely inhibitory couplings
(van Vreeswijk et al., 1994). In order to get a response at all in such a system, we
need a constant stimulus I0 or, equivalently, a negative firing threshold
![]() < 0. The stability criterion, however, is equivalent to that of the
previous example.
< 0. The stability criterion, however, is equivalent to that of the
previous example.
Figure 8.6 summarizes the stability arguments analogously to Fig. 8.5. In Fig. 8.6A all neurons have fired synchronously at t = 0 and do so again at t = T when the inhibitory postsynaptic potential has decayed so that the threshold condition,
| (8.17) |
![\begin{minipage}[t]{0.45\textwidth}
{\bf A}\\
\par\includegraphics[width=\text...
...raphics[width=\textwidth]{Figs-ch-oscillations/lock_inhib_b.eps}
\end{minipage}](img1322.gif) |
We consider a homogeneous populations of SRM neurons that are close to a
periodic state of synchronized activity. We assume that the population
activity in the past consists of a sequence of rectangular pulses as specified
in Eq. (8.11). We determine the period T and the sequence of
half-widths ![]() of the rectangular pulses in a self-consistent manner.
In order to prove stability, we need to show that the amplitude A(k T)
increases while the halfwidth
of the rectangular pulses in a self-consistent manner.
In order to prove stability, we need to show that the amplitude A(k T)
increases while the halfwidth ![]() decreases as a function of k. To
do so we start from the noise-free population equation (7.13) that we
recall here for convenience
decreases as a function of k. To
do so we start from the noise-free population equation (7.13) that we
recall here for convenience
As a first step, we calculate the potential
hPSP(t|![]() ). Given
hPSP we can find the period T from the threshold condition and
also the derivatives
). Given
hPSP we can find the period T from the threshold condition and
also the derivatives
![]() h and
h and
![]() h required for
Eq. (7.13). In order to obtain
hPSP, we substitute
Eq. (8.11) in
(6.8), assume
h required for
Eq. (7.13). In order to obtain
hPSP, we substitute
Eq. (8.11) in
(6.8), assume
![]()
![]() T, and integrate. To first order in
T, and integrate. To first order in
![]() we obtain
we obtain
In the second step we determine the period T. To do so, we consider a
neuron in the center of the square pulse which has fired its last spike
at ![]() = 0. Since we consider noiseless neurons the relative order of
firing of the neurons cannot change. Consistency of the ansatz (8.11)
thus requires that the next spike of this neuron must occur at t = T, viz. in
the center of the next square pulse. We use
= 0. Since we consider noiseless neurons the relative order of
firing of the neurons cannot change. Consistency of the ansatz (8.11)
thus requires that the next spike of this neuron must occur at t = T, viz. in
the center of the next square pulse. We use ![]() = 0 in the threshold
condition for spike firing which yields
= 0 in the threshold
condition for spike firing which yields
In the population equation (8.18) we need the derivative of hPSP,
We now apply Eq. (8.21) to a population of
SRM0 neurons.
For SRM0 neurons we have
![]() (x, s) =
(x, s) = ![]() (s), hence
(s), hence
![]() h = 0 and
hPSP(t|
h = 0 and
hPSP(t|![]() ) = h(t) = J0
) = h(t) = J0![]()
![]() (t + k T). For a standard
(t + k T). For a standard ![]() kernel (e.g. an
exponentially decaying function), we have
kernel (e.g. an
exponentially decaying function), we have
![]() (T) > 0 whatever T and thus
(T) > 0 whatever T and thus
![\vbox{
\vspace{5mm}
\centerline{
\includegraphics[height=15mm,width=60mm]{Figs...
...egraphics[height=12mm,width=60mm]{Figs-ch-oscillations/pout.dat.square.ioft}
}}](img1335.gif) |
The growth of amplitude corresponds to a compression of the width of the
pulse. It can be shown that the `corner neurons' which have fired at time
±![]() fire their next spike at
T±
fire their next spike at
T±![]() where
where
![]() =
= ![]() A(0)/A(T).
Thus the square pulse remains normalized
as it should be. By iteration of the argument for t = k T with
k = 2, 3, 4,... we see that the sequence
A(0)/A(T).
Thus the square pulse remains normalized
as it should be. By iteration of the argument for t = k T with
k = 2, 3, 4,... we see that the sequence ![]() converges to zero and the square pulses approach a Dirac
converges to zero and the square pulses approach a Dirac ![]() -pulse under
the condition that
h'(T) =
-pulse under
the condition that
h'(T) = ![]()
![]() (k T) > 0. In other words, the
T-periodic synchronized solution with T given by Eq. (8.20) is stable,
if the input potential h at the moment of firing is rising
(Gerstner et al., 1996b).
(k T) > 0. In other words, the
T-periodic synchronized solution with T given by Eq. (8.20) is stable,
if the input potential h at the moment of firing is rising
(Gerstner et al., 1996b).
In order for the sequence of square pulses to be an exact solution of the
population equation, we must require that the factor
in the square brackets of Eq. (8.18)
remains constant over the width of a pulse. The derivatives of
Eq. (8.19), however, do depend on t.
As a consequence, the form of the
pulse changes over time as is visible in Fig. 8.7. The activity as
a function of time was obtained by a numerical integration of the population
equation with a square pulse as initial condition for a network of SRM0
neurons coupled via (8.10) with weak inhibitory coupling J = - 0.1 and
delay
![]() = 2ms. For this set of parameters h' > 0 and locking
is possible.
= 2ms. For this set of parameters h' > 0 and locking
is possible.
The framework of the population equation allows us also to extend the locking argument to noisy SRM0 neurons. At each cycle, the pulse of synchronous activity is compressed due to locking if h'(T) > 0. At the same time it is smeared out because of noise. To illustrate this idea we consider SRM0 neurons with Gaussian noise in the reset.
In the case of noisy reset, the interval distribution can
be written as
PI(t|![]() ) =
) = ![]() dr
dr ![]() [t -
[t - ![]() - T(
- T(![]() , r)]
, r)] ![]()
![]() (r);
cf. Eq. (5.68). We insert the
interval distribution into
the population equation
A(t) =
(r);
cf. Eq. (5.68). We insert the
interval distribution into
the population equation
A(t) = ![]() PI(t|
PI(t|![]() ) A(
) A(![]() ) d
) d![]() and find
and find
We now search for periodic solutions.
As shown below, a limit cycle solution of
Eq. (8.24) consisting of a
sequence of Gaussian pulses exists if the noise amplitude ![]() is small
and
(h'/
is small
and
(h'/![]() ) > 0. The width d of the activity pulses
in the limit cycle is proportional to the noise level
) > 0. The width d of the activity pulses
in the limit cycle is proportional to the noise level ![]() .
A simulation of locking in the presence of
noise is shown in Fig. 8.8. The network of SRM0 neurons has
inhibitory connections (J0 = - 1) and is coupled via the response kernel
(8.10) with a transmission delay of
.
A simulation of locking in the presence of
noise is shown in Fig. 8.8. The network of SRM0 neurons has
inhibitory connections (J0 = - 1) and is coupled via the response kernel
(8.10) with a transmission delay of
![]() = 2 ms.
Doubling the noise level
= 2 ms.
Doubling the noise level ![]() leads to activity pulses with twice the
width.
leads to activity pulses with twice the
width.
![\begin{minipage}[t]{0.45\textwidth}
{\bf A}\\
\par\includegraphics[width=\text...
...\
\par\includegraphics[width=\textwidth]{pout.dat.lock.s05.eps}
\end{minipage}](img1338.gif) |
In order to calculate the width of the activity pulses in a locked state, we
look for periodic pulse-type solutions of Eq. (8.24). We
assume that the pulses are Gaussians with width d and repeat with period
T, viz.,
A(t) = ![]()
![]() d(t - k T). The pulse width d will be
determined self-consistently from Eq. (8.24). The integral over r in
Eq. (8.24) can be performed and yields a Gaussian with width
d(t - k T). The pulse width d will be
determined self-consistently from Eq. (8.24). The integral over r in
Eq. (8.24) can be performed and yields a Gaussian with width
![]() = [d2 +
= [d2 + ![]() ]1/2. Equation (8.24) becomes
]1/2. Equation (8.24) becomes
Let us work out the self-consistency condition and focus on the pulse around
t ![]() 0. It corresponds to the k = 0 term on the left-hand side which
must equal the k = - 1 term on the right-hand side of
Eq. (8.25). We
assume that the pulse width is small d
0. It corresponds to the k = 0 term on the left-hand side which
must equal the k = - 1 term on the right-hand side of
Eq. (8.25). We
assume that the pulse width is small d ![]() T and expand Tb(t) to linear
order around
Tb(0) = T. This yields
T and expand Tb(t) to linear
order around
Tb(0) = T. This yields
We have seen that, on the one hand, the state of asynchronous firing is typically unstable for low levels of noise. On the other hand, the fully locked state may be unstable as well if transmission delay and length of the refractory period do not allow spikes to be triggered during the rising phase of the input potential. The natural question is thus: What does the network activity look like if both the asynchronous and the fully locked state are unstable?
![\begin{minipage}[t]{0.38\textwidth}
{\bf A}\\
\par\includegraphics[width=\text...
...phics[width=\textwidth]{Figs-ch-oscillations/lock_cluster_b.eps}
\end{minipage}](img1358.gif) |
Figure 8.9A shows an example of an excitatory network with vanishing transmission delay and a rather long refractory period as compared to the rising phase of the postsynaptic potential. As a consequence, the threshold condition is met when the postsynaptic potential has already passed its maximum. The fully locked state is thus unstable. This, however, does not mean that the network will switch into the asynchronous mode. Instead, the neurons may split into several subgroups (``cluster'') that fire alternatingly. Neurons within each group stay synchronized. An example of such a cluster state with two subgroups is illustrated in Fig. 8.9B. Action potentials produced by neurons from group 1 trigger group 2 neurons and vice versa. The population activity thus oscillates with twice the frequency of an individual neuron.
In general, there is an infinite number of different cluster states that can be indexed by the number of subgroups. The length T of the inter-spike interval for a single neuron and the number of subgroups n in a cluster state are related by the threshold condition for spike triggering (Kistler and van Hemmen, 1999; Chow, 1998),
| (8.29) |
| (8.30) |
In Section 8.1 we have seen that the state of asynchronous firing in a SRM network is always unstable in the absence of noise. We now see that even if the fully locked state is unstable the network is not firing asynchronously but usually gets stuck in one of many possible cluster states. Asynchronous firing can only be reached asymptotically by increasing the number of subgroups so as to ``distribute'' the spike activity more evenly in time. Individual neurons, however, will always fire in a periodical manner. Nevertheless, increasing the number of subgroups will also reduce the amplitude of the oscillations in the input potential and the firing time of the neurons becomes more and more sensitive to noise. The above statement that asynchrony can only be reached asymptotically is therefore only valid in strictly noiseless networks.
A final remark on the stability of the clusters is in order. Depending on the form of the postsynaptic potential, the stability of the locked state may be asymmetric in the sense that neurons that fire too late are pulled back into their cluster, neurons that have fired to early, however, are attracted by the cluster that has just fired before. If the noise level is not too low, there are always some neurons that drop out of their cluster and drift slowly towards an adjacent cluster (Ernst et al., 1995; van Vreeswijk, 1996).
To illustrate the relation between the instability of the state of
asynchronous firing and cluster states, we return to the network of SRM0
neurons with noisy reset that we have studied in Section 8.1. For low noise (
![]() = 0.04), the asynchronous firing
state is unstable whatever the axonal transmission delay;
cf. Fig. 8.2. With an axonal delay of 2ms, asynchronous firing
is unstable with respect to an oscillation with
= 0.04), the asynchronous firing
state is unstable whatever the axonal transmission delay;
cf. Fig. 8.2. With an axonal delay of 2ms, asynchronous firing
is unstable with respect to an oscillation with ![]() . The population
splits into 3 different groups of neurons that fire with a period of about
8ms. The population activity, however, oscillates with a period of 2.7ms;
cf. Fig. 8.10A. With a delay of 1.2ms, the asynchronous
firing state has an instability with respect to
. The population
splits into 3 different groups of neurons that fire with a period of about
8ms. The population activity, however, oscillates with a period of 2.7ms;
cf. Fig. 8.10A. With a delay of 1.2ms, the asynchronous
firing state has an instability with respect to ![]() so that the
population activity oscillates with a period of about 1.6ms. The population
splits into 5 diferent groups of neurons that fire with a period of about
8ms; cf. Fig. 8.10B.
so that the
population activity oscillates with a period of about 1.6ms. The population
splits into 5 diferent groups of neurons that fire with a period of about
8ms; cf. Fig. 8.10B.
![\begin{minipage}{0.40\textwidth}
{\bf A}
\par\includegraphics[width=\textwidth]...
....2s.eps}
\includegraphics[width=\textwidth]{sp-train-d1.2s.eps}
\end{minipage}](img1363.gif) |
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
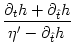
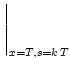 .
.