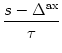In Section 6.4 and throughout Chapter 7, we have assumed that the network is in a state of asynchronous firing. In this section, we study whether asynchronous firing can indeed be a stable state of a fully connected population of spiking neurons - or whether the connectivity drives the network towards oscillations. For the sake of simplicity, we restrict the analysis to SRM0 neurons; the same methods can, however, be applied to integrate-and-fire neurons or general SRM neurons.
For SRM0 neurons, the membrane potential is given by
| ui(t) = |
(8.1) |
The state of asynchronous firing corresponds to a fixed point A(t) = A0 of the population activity. We have already seen in Chapter 6.4 how the fixed point A0 can be determined either numerically or graphically. To analyze its stability we assume that for t > 0 the activity is subject to a small perturbation,
The perturbation of the potential causes some neurons to fire earlier (when
the change in h is positive) others to fire later (whenever the change is
negative). The perturbation may therefore build up (![]() > 0, the
asynchronous state is unstable) or decay back to zero (
> 0, the
asynchronous state is unstable) or decay back to zero (![]() < 0, the
asynchronous state is stable). At the transition between the region of
stability and instability the amplitude of the perturbation remains constant
(
< 0, the
asynchronous state is stable). At the transition between the region of
stability and instability the amplitude of the perturbation remains constant
(![]() = 0, marginal stability of the asynchronous state). These transition
points, defined by
= 0, marginal stability of the asynchronous state). These transition
points, defined by ![]() = 0, are determined now.
= 0, are determined now.
We start from the population integral equation
A(t) = ![]() PI(t|
PI(t|![]() ) A(
) A(![]() ) d
) d![]() that has been introduced in Chapter 6.3.
Here
PI(t|
that has been introduced in Chapter 6.3.
Here
PI(t|![]() ) is the input-dependent interval
distribution, i.e., the probability density of emitting a spike
at time t given that the last spike occured at time
) is the input-dependent interval
distribution, i.e., the probability density of emitting a spike
at time t given that the last spike occured at time ![]() .
We have seen in Chapter 7 that the linearized population
activity equation can be written in the form
.
We have seen in Chapter 7 that the linearized population
activity equation can be written in the form
We have written Eq. (8.8) as a combination of two
requirements, i.e., an amplitude condition
Sf(![]() ) = 1 and a phase condition
) = 1 and a phase condition
![]() (
(![]() ) mod 2
) mod 2![]() = 0. Let us discuss the
general structure of the two conditions. First, if
Sf(
= 0. Let us discuss the
general structure of the two conditions. First, if
Sf(![]() )
)![]() 1 for
all frequencies
1 for
all frequencies ![]() , an oscillatory perturbation cannot build up. All
oscillations decay and the state of asynchronous firing is stable. We conclude
from Eq. (8.7) that by increasing the absolute value | J0| of the
coupling constant, it is always possible to increase
Sf(
, an oscillatory perturbation cannot build up. All
oscillations decay and the state of asynchronous firing is stable. We conclude
from Eq. (8.7) that by increasing the absolute value | J0| of the
coupling constant, it is always possible to increase
Sf(![]() ). The
amplitude condition can thus be met if the excitatory or inhibitory feedback
from other neurons in the population is sufficiently strong. Second, for a
bifurcation to occur we need in addition that the phase condition is
met. Loosely speaking, the phase condition implies that the feedback from
other neurons in the network must arrive just in time to keep the oscillation
going. Thus the axonal signal transmission time and the rise time of the
postsynaptic potential play a critical role during oscillatory activity
(Mattia and Del Giudice, 2001; Treves, 1993; Abbott and van Vreeswijk, 1993; Brunel, 2000; Neltner et al., 2000; Gerstner, 1995; Vreeswijk, 2000; Gerstner, 2000b; Brunel and Hakim, 1999; Ernst et al., 1995; Gerstner and van Hemmen, 1993; Tsodyks et al., 1993).
). The
amplitude condition can thus be met if the excitatory or inhibitory feedback
from other neurons in the population is sufficiently strong. Second, for a
bifurcation to occur we need in addition that the phase condition is
met. Loosely speaking, the phase condition implies that the feedback from
other neurons in the network must arrive just in time to keep the oscillation
going. Thus the axonal signal transmission time and the rise time of the
postsynaptic potential play a critical role during oscillatory activity
(Mattia and Del Giudice, 2001; Treves, 1993; Abbott and van Vreeswijk, 1993; Brunel, 2000; Neltner et al., 2000; Gerstner, 1995; Vreeswijk, 2000; Gerstner, 2000b; Brunel and Hakim, 1999; Ernst et al., 1995; Gerstner and van Hemmen, 1993; Tsodyks et al., 1993).
Let us apply the above results to SRM0 neurons with noise in the reset. We
assume that neurons are in a state of asynchronous firing with activity A0.
As we have seen in Chapter 5, the interval distribution for noisy
reset is a Gaussian centered at
T0 = 1/A0. The filter function
![]() is a
is a
![]() -function,
-function,
![]() (x) =
(x) = ![]() (x)/
(x)/![]() ; cf. Table 7.1.
Hence Eq. (8.7) is of the form
; cf. Table 7.1.
Hence Eq. (8.7) is of the form
In order to analyze Eq. (8.9) numerically we have to specify the response kernel. For the sake of simplicity we choose a delayed alpha function,
Figure 8.1 shows Sf as a function of
![]() T0. Since
Sf = 1 is a necessary condition for a bifurcation, it is apparent that
bifurcations can occur only for frequencies
T0. Since
Sf = 1 is a necessary condition for a bifurcation, it is apparent that
bifurcations can occur only for frequencies
![]()
![]()
![]() = n 2
= n 2![]() /T0 with integer n where
T0 = 1/A0 is the typical inter-spike
interval. We also see that higher harmonics are only relevant for low levels
of noise. For
/T0 with integer n where
T0 = 1/A0 is the typical inter-spike
interval. We also see that higher harmonics are only relevant for low levels
of noise. For
![]()
![]() 0 the absolute value of the denominator of
(8.9) is
2| sin(
0 the absolute value of the denominator of
(8.9) is
2| sin(![]() T0/2)| and bifurcations can occur for all
higher harmonics. At a high noise level, however, the asynchronous state is
stable even with respect to perturbations at
T0/2)| and bifurcations can occur for all
higher harmonics. At a high noise level, however, the asynchronous state is
stable even with respect to perturbations at
![]()
![]()
![]() .
.
![\hbox{
\hspace{20mm}
\includegraphics[width=50mm,height=25mm]{Figs-ch-oscillations/pout.dat.feedback}
}
\vspace{1mm}](img1299.gif) |
A bifurcation at
![]()
![]()
![]() implies that the period of the
perturbation is identical to the firing period of individual neurons. Higher
harmonics correspond to instabilities of the asynchronous state towards
cluster states (Kistler and van Hemmen, 1999; Ernst et al., 1995; Gerstner and van Hemmen, 1993; Golomb et al., 1992; Golomb and Rinzel, 1994): each
neuron fires with a mean period of T0, but the population of neurons splits
up in several groups that fire alternatingly so that the overall activity
oscillates several times faster; cf. Section 8.2.3.
implies that the period of the
perturbation is identical to the firing period of individual neurons. Higher
harmonics correspond to instabilities of the asynchronous state towards
cluster states (Kistler and van Hemmen, 1999; Ernst et al., 1995; Gerstner and van Hemmen, 1993; Golomb et al., 1992; Golomb and Rinzel, 1994): each
neuron fires with a mean period of T0, but the population of neurons splits
up in several groups that fire alternatingly so that the overall activity
oscillates several times faster; cf. Section 8.2.3.
Figure 8.1 illustrates the amplitude condition for the solution of
Eq. (8.9). The numerical solutions of the full equation
(8.9) for different values of the delay
![]() and
different levels of the noise
and
different levels of the noise ![]() are shown in the bifurcation diagram of
Fig. 8.2. The insets show simulations that illustrate the behavior of
the network at certain combinations of transmission delay and noise level.
are shown in the bifurcation diagram of
Fig. 8.2. The insets show simulations that illustrate the behavior of
the network at certain combinations of transmission delay and noise level.
Let us consider for example a network with transmission delay
![]() = 2 ms, corresponding to a x-value of
= 2 ms, corresponding to a x-value of
![]() /T0 = 0.25 in
Fig. 8.2. The phase diagram predicts that, at a noise level of
/T0 = 0.25 in
Fig. 8.2. The phase diagram predicts that, at a noise level of
![]() = 0.5 ms, the network is in a state of asynchronous firing. The
simulation shown in the inset in the upper right-hand
corner confirms that the activity fluctuates around a constant value of
A0 = 1/T0 = 0.125 kHz.
= 0.5 ms, the network is in a state of asynchronous firing. The
simulation shown in the inset in the upper right-hand
corner confirms that the activity fluctuates around a constant value of
A0 = 1/T0 = 0.125 kHz.
If the noise level of the network is significantly reduced, the system crosses
the short-dashed line. This line is the boundary at which the constant
activity state becomes unstable with respect to an oscillation with
![]()
![]() 3 (2
3 (2![]() /T0). Accordingly, a network simulation with a noise
level of
/T0). Accordingly, a network simulation with a noise
level of
![]() = 0.1 exhibits an oscillation of the population activity
with period
Tosc
= 0.1 exhibits an oscillation of the population activity
with period
Tosc ![]() T0/3
T0/3 ![]() 2.6 ms.
2.6 ms.
Keeping the noise level constant but reducing the transmission delay
corresponds to a horizontal move across the phase diagram in
Fig. 8.2. At some point, the system crosses the solid line that marks
the transition to an instability with frequency
![]() = 2
= 2![]() /T0. Again,
this is confirmed by a simulation shown in the inset in the upper
left corner. If we now decrease the noise level, the oscillation becomes even
more pronounced (bottom right).
/T0. Again,
this is confirmed by a simulation shown in the inset in the upper
left corner. If we now decrease the noise level, the oscillation becomes even
more pronounced (bottom right).
![\hbox{
\hspace{10mm}
\includegraphics[width=100mm,height=89mm]{Figs-ch-oscillations/pout.dat.plot-all.eps}
}
\vspace{8mm}](img1300.gif) |
In the limit of low noise, the asynchronous network state is unstable for
virtually all values of the delay. The region of the phase diagram in
Fig. 8.2 around
![]() /T0
/T0 ![]() 0.1 which looks
stable hides instabilities with respect to the higher harmonics
0.1 which looks
stable hides instabilities with respect to the higher harmonics ![]() and
and ![]() which are not shown. We emphasize that the specific location
of the stability borders depends on the form of the postsynaptic response
function
which are not shown. We emphasize that the specific location
of the stability borders depends on the form of the postsynaptic response
function ![]() . The qualitative features of the phase diagram in
Fig. 8.2 are generic and hold for all kinds of response kernels.
. The qualitative features of the phase diagram in
Fig. 8.2 are generic and hold for all kinds of response kernels.
The numerical results apply to the response kernel
![]() (s) defined in
(8.10) which corresponds to a synaptic current
(s) defined in
(8.10) which corresponds to a synaptic current ![]() (s) with zero
rise time; cf. (4.2) and (4.34). What happens if
(s) with zero
rise time; cf. (4.2) and (4.34). What happens if ![]() is a
double exponential with rise time
is a
double exponential with rise time
![]() and decay time
and decay time
![]() ? In this case, the right-hand side of (8.9) has an
additional factor
[1 + i
? In this case, the right-hand side of (8.9) has an
additional factor
[1 + i ![]()
![]() ]-1 which leads to two
changes. First, due to the reduced amplitude of the feedback, instabilities
with frequencies
]-1 which leads to two
changes. First, due to the reduced amplitude of the feedback, instabilities
with frequencies
![]() >
> ![]() are suppressed. The tongues
for the higher harmonics are therefore smaller. Second, the phase of the
feedback changes. Thus all tongues of frequency
are suppressed. The tongues
for the higher harmonics are therefore smaller. Second, the phase of the
feedback changes. Thus all tongues of frequency ![]() are moved
horizontally along the x-axis by an amount
are moved
horizontally along the x-axis by an amount
![]() /T0 = - arctan(
/T0 = - arctan(![]()
![]() )/(n 2
)/(n 2![]() ).
).
What happens if the excitatory interaction is replaced by inhibitory coupling?
A change in the sign of the interaction corresponds to a phase shift of ![]() .
For each harmonic, the region along the delay axis where the asynchronous state
is unstable for excitatory coupling (cf. Fig. 8.2)
becomes stable for inhibition and vice versa. In other words, we
simply have to shift the instability tongues for each frequency
.
For each harmonic, the region along the delay axis where the asynchronous state
is unstable for excitatory coupling (cf. Fig. 8.2)
becomes stable for inhibition and vice versa. In other words, we
simply have to shift the instability tongues for each frequency ![]() horizontally by an amount
horizontally by an amount
![]() /T0 = 1/(2n). Apart from that the pattern
remains the same.
/T0 = 1/(2n). Apart from that the pattern
remains the same.
![\centerline{
\includegraphics[width=8cm]{brunel-3b-edit.eps}
}](img1309.gif) |
Our discussion of random-connectivity networks in Chapter 6.4.3 has been focused on the stationary state of asynchronous firing. The stability analysis of the asynchronous state in such randomly connected networks is completely analogous to the approach sketched in Eqs. (8.3) - (8.6) except that the linearization is performed on the level of the density equations (Brunel, 2000; Brunel and Hakim, 1999). Close to the asynchronous state, the activity can be written as A(t) = A0 + A1(t) and the membrane potential distribution as p(u, t) = p0(u) + p1(u, t). Here p0(u) is the stationary distribution of membrane potential in the state of asynchronous firing [cf. Eqs. (6.27) and (6.28)] and p1(u, t) is a small time-dependent perturbation. The stability analysis requires a linearization of the Fokker-Planck equation (6.21) with respect to p1 and A1.
For short transmission delays, the asynchronous
state
A(t) ![]() A0
can loose its stability towards an oscillation
with a frequency that is much faster
than the single-neuron firing rate.
Brunel (2000)
distinguishes two different variants
of such fast oscillations.
First, as in the previous example
there are cluster states where
the neuronal population splits into a few subgroups.
Each neuron fires nearly regularly
and within a cluster neurons are almost fully
synchronized; cf. Section 8.2.3.
Second, there are synchronous irregular
states where the global activity oscillates
while individual neurons have
a broad distribution of interspike intervals;
cf. Fig. 8.3.
We will come back to synchronous irregular
states in Section 8.3.
A0
can loose its stability towards an oscillation
with a frequency that is much faster
than the single-neuron firing rate.
Brunel (2000)
distinguishes two different variants
of such fast oscillations.
First, as in the previous example
there are cluster states where
the neuronal population splits into a few subgroups.
Each neuron fires nearly regularly
and within a cluster neurons are almost fully
synchronized; cf. Section 8.2.3.
Second, there are synchronous irregular
states where the global activity oscillates
while individual neurons have
a broad distribution of interspike intervals;
cf. Fig. 8.3.
We will come back to synchronous irregular
states in Section 8.3.
© Cambridge University Press
This book is in copyright. No reproduction of any part
of it may take place without the written permission
of Cambridge University Press.
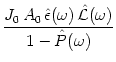 = Sf(
= Sf(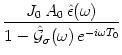 = Sf(
= Sf( exp
exp